Как сделать сигну в вк фото
Что такое сигна в ВК?
Вконтакте существует большое количество фейковых страниц (см. купить страницу Вконтакте). Если для пользователя важно подтвердить, что это его настоящая страница, используется несколько механизмов.
Один из них — подтверждение страницы (см. как получить галочку вконтакте). Делается он через официальный запрос в службу поддержки.
Вариант проще — это получение и публикация сигны. В данной инструкции я попытаюсь объяснить вам, что такое сигна в ВК, и как ее сделать.
Оглавление:
Что такое сигна Вконтакте
Это фотография, на которой изображено ваше id (см. как узнать свой id вконтакте), в любом удобном варианте. Самый простой вариант — вы пишите на листке бумаги свой id, и фотографируетесь с ним (см. как поменять id вконтакте). Вот пример.
Таким образом вы подтверждаете, что вы реальный человек, и эта страница принадлежит вам.
Сейчас очень модно получать сигны со всевозможными подписями. Имя, название группы и т.д. На заказ вам могут сделать любую сигну.
Имя, название группы и т.д. На заказ вам могут сделать любую сигну.
Как сделать сигну в ВК
Ну с самостоятельным вариантом все понятно. А если нужно, чтобы ее сделал другой человек?
Вам нужно найти группу, где люди предлагают создание сигн за деньги. Где их найти (см. поиск групп вк)? Идем в раздел «Группы», и в поиске пишем «Сделай мне сигну». Фильтруем по сообществам, и в результатах поиска находим подходящую группу. Переходим в нее.
На стене группы люди пишут объявление, с предложением сделать сигну. Найдите подходящий вариант. Можно даже получить ее совершенно бесплатно.
Свяжитесь с человеком, и договоритесь об условиях.
Видео урок: что означает сигна в ВК
Читайте также:
Если вам важно, чтобы другие пользователи были уверены в надежности вашей страницы, то можете рассматривать вариант использования сигн.
Мой мир
Вконтакте
Одноклассники
Google+
(1 участников, средняя оценка: 5,00 из 5) Загрузка. ..
..Что такое сигна в вк, как сделать сигну вконтакте и на этом заработать | Блог Артема Полуэктова
(2 голосов, оценка: 5,00 из 5) Загрузка… Автор: Admin Опубликовано: 2016-06-23в5:00
Вы знаете, что такое сигна в вк? А как сделать сигну вконтакте? В последнее время делать сигну стало очень популярным. Она стала все чаще появляться в социальных сетях.
Всем привет! Из этой статьи вы узнаете о сигне. В частности что такое сигна, если вас заинтересует, я вам покажу как сделать сигну. Знаете в 21-ом веке уже никого ничем не удивишь, добрались до того, что ее делают прямо на своем теле! Есть и такие, кто предлагает другому сделать у себя ее за деньги. А для кого то это бизнес, а для кого то и работа. Давайте обо всем по порядку.
Что такое это, и что такое Signa в вк?
Что это такое? Сигна, дословно с английского переводиться как знак. Изначально к нам это пришло с запада. Смысл был таков, человек брал листок бумаги, на нем писал адрес своей страницы от социальной сети и фотографировал себя. Затем выкладывал эту фотографию на своей странице. Догадываетесь для чего так делали люди?)
Затем выкладывал эту фотографию на своей странице. Догадываетесь для чего так делали люди?)
Этим действием человек подтверждал, что аккаунт его.
Классная статья: Как поменять пароль в контактеРазумеется на этом никто не остановился. Далее было придумано писать уже не на листке бумаги или фотографии, а непосредственно на своем теле. Что писали? Сначала, люди просто хотели где нибудь «засветиться,» чтобы на них обратили внимание. Или писали имя поклонника или поклонницы. Так как сейчас социальные сети в наше время являются неотъемлемой частью ее распространение дошло и до них, самую известную соц. сеть в России вконтакте. Сейчас это превратилась в большой бизнес. Кстати некоторые на этом не плохо зарабатывают). Заинтересовало?) Тогда рассказываю далее. Как сделать Signe?
Как сделать сигну и на этом заработать Вконтакте (в вк) ?
Как я уже сказал сигна — это уже целая бизнес индустрия. Как она делается, или как люди научились на этом зарабатывать? По сути сигну превратили просто в рекламу.
Есть два варианта зарабатывания денег на ней.
1. Берется просто листок бумаги на нем она пишется, т.е. реклама и вы с ней фотографируетесь, так как сейчас большое распространение получил именно онлайн бизнес, обычно на листке бумаги пишут адрес сайта или популярной группы вк. Этот вариант, предлагают обычно известным или популярным людям.
Разумеется, как и любая реклама, такие действия оплачиваются рекламодателем.
Классная статья: Кто такой фейк в контакте, как создать, распознать, поиск2. Сигна пишется на своем теле. Обычно на пикантных местах, груди и попе. Или просто на листке бумаги, а человек нижнем белье держа ее фотографируется с ней. Если исходить из такого варианта, уже любой желающий может делать такого вида рекламу, очень востребована такая сигна в вк (вконтакте), правда для этого вам нужно договориться с рекламодателем. Кстати, вы и сами можете убедиться, что эта индустрия заработка востребована вконтакте.
Для этого достаточно вбить в поиске групп в вк «сигна. »
»
Да, вы наверное знаете, что администрация контакта блокирует аккаунты на право и налево. По поводу сигн, можете особо не переживать, если это не откровенная эротика, администрация ровно дышит к таким фотографиям.
Лучше видео, которое я нашел на тему о сигнах.
На этом на сегодня все. Всем удачи в новых начинаниях! Интересные статьи для Вас:
Что такое сигна — зачем и как её делают, примеры с фото
Чтобы подтвердить авторство фото или видео вконтакте, необходимо отчетливо понимать, что такое сигна, для чего, когда ее используют. Этот термин относительно молодой, изначально появился у пользователей социальных сетей, считается хорошей возможностью сократить количество фейков. Для современной молодежи стал оригинальным знаком благодарности, способом выражения искренности романтических чувств. Поэтому всем пользователям сети не помешает выяснить, что такое сигны в ВК, как правильно пользоваться таким атрибутом.
Что значит сделать сигну
По определенному электронному адресу можно найти страницу пользователя вконтакте и другой социальной сети. Чтобы лишний раз удостовериться, что собеседник связался с вашим аккаунтом, он может попросить подтверждение подлинности контента. Вариантов несколько, и один из них – сигна с именем на фотографиях. Реальные фото выкладывают на аватарках или по запросу того или иного пользователя. Такими изображениями пестрят социальные сети, они в неограниченном количестве блуждают по интернету.
Значение слова sign в переводе с английского языка звучит примерно так: «обозначающее подпись». Грубо говоря, это фотография-подарок, как вид благодарности в социальных сетях. Сигны могут набрать много голосов, стать предметом публичности или быть отправлены личным сообщением, как что-то сокровенное адресату. Изначально, на чистом листке бумаги пользователи писали адрес своей страницы социальной сети и фотографировались, чтобы исключить подозрение на фейк.
Позднее сигны стали подарком, сюрпризом, романтическим признанием. Таким знаком внимания часто делится современная молодежь, а тематическую надпись оставляет не только на бумаге, но и на одной из видимых частей тела. Это тоже имеет значение, поскольку таким способом можно подтвердить не только подлинность аккаунта ВК, но и собственной личности. Так делают сигны на руке, ноге, груди, лице, других частях тела (по своему желанию).
Зачем нужны сигны
Перед тем как сделать сигну в ВК, важно не только понимать значение этого молодежного сленга, но и его необходимость на практике. Некоторые пользователи загадочную сигну делают названием страницы, чтобы привлечь большое количество поклонников своей таинственностью. Это фишка красивых девушек, которые привлекают внимание к своей фигуре, но изначально закрывают лицо сигной. Так можно стать кумиром современной молодежи.
Второй вариант использования сигны, когда хотят узнать, симпатичные девушки реальные или фейк, блуждающий на просторах интернета. Например, просят на листе бумаги или части тела написать имя парня, выполнить селфи с пересылкой адресату. Это касается и спортивных, привлекательных парней, которые стали частыми посетителями соцсетей. Некоторые девушки, особенно любопытные и подозрительные, хотят выяснить, их кумир – фейк или настоящий. К тому же, делать сигны считается модным молодежным течением, трендом современной молодежи.
Например, просят на листе бумаги или части тела написать имя парня, выполнить селфи с пересылкой адресату. Это касается и спортивных, привлекательных парней, которые стали частыми посетителями соцсетей. Некоторые девушки, особенно любопытные и подозрительные, хотят выяснить, их кумир – фейк или настоящий. К тому же, делать сигны считается модным молодежным течением, трендом современной молодежи.
Что можно написать
За последний год загадочная сигна пользуется высокой популярностью, и многие страницы вконтакте не обходятся без ее непосредственного участия. На таких реалистичных фото различаются позы и надписи, цели выполнения селфи и отзывы пользователей социальных сетей. Снимки могут вызывать повышенный интерес общественности своим своеобразным видом, и тогда обязательно наберут много лайков, положительных комментариев, что особенно актуально для лайфхак.
Изучая, что такое сигна, возникает вопрос, какая подпись должна быть и уместна на снимке. Вариантов существует много, например, что попросят сделать адресата, то и написать на листе бумаги, частях тела. Чаще девушки оставляют признания в любви на груди и руках или просто пишут свое имя, имя конкретного парня для подтверждения подлинности страницы, паблика. Главное – фотографируйтесь оригинально и от души, такие снимки обязательно не останутся без внимания. Вместо надписей можно использовать рисунки, например, с тематических сайтов и не только.
Чаще девушки оставляют признания в любви на груди и руках или просто пишут свое имя, имя конкретного парня для подтверждения подлинности страницы, паблика. Главное – фотографируйтесь оригинально и от души, такие снимки обязательно не останутся без внимания. Вместо надписей можно использовать рисунки, например, с тематических сайтов и не только.
Что нарисовать
Графические изображения тоже являются частью актуальной темы современной молодежи, что же такое сигна. Изначально все начиналось с подписи имени, но со временем такая фотография-подарок приобрела более глубокий смысл, емкое содержание. Например, сентиментальные девушки в рисунках на своем теле оставляли романтическое послание понравившимся парням. Другие пользователи социальных сетей и вовсе создавали на теле истинные шедевры для повышения количества лайков, комментариев.
Во всемирной паутине имеются специальные сайты, посвященные теме создания сигны с переводами и без. Там же можно найти ценные рекомендации, как проверить такой снимок на подлинность, исключить очередную подделку в сети. Даже некоторые знаменитости при виртуальном общении с фанатами и поклонниками вместо личного автографа используют сигны. Таким же способом известные люди подтверждают подлинность электронных адресов в социальных сетях. Полезно знать, как правильно делать сигны, и какие хитрости этого увлекательного процесса существуют
Видео-сигны
Сигны стали не только хорошей возможностью подтвердить свою личность в социальных сетях, но и выгодным источником получения дохода. Особенно популярные пользователи с огромным числом подписчиков продают собственные фото с подписями своих фанатов за деньги. Этим промышляют и многие «начинающие знаменитости», но делают это еще в качестве рекламы, пиара.
За последнее время среди молодежи востребованными стали не только фото-снимки, но и непродолжительные (короткометражные) видео-ролики, тоже подтверждающие личность автора, передающие определенное послание со смыслом адресату. Если говорить о классическом видео-сигне, ниже представлена домашняя инструкция по созданию такого оригинального матерела:
- Создать на листе бумаги или руке тематическую надпись, которую попросил для подтверждения адресат.

- Выполнить видео-ролик, переслать его адресату. Проще говоря, в видео человек показывает надпись с именем друга, подруги и т.д.
- Отправить готовую видео-работу на стену или личным сообщением – на усмотрение создателя ролика.
На руке
Это самый популярный вариант для соцсетей. Для создания сигна на руке необходимо действовать в следующей последовательности:
- Написать на оголенной руке надпись, которую попросили.
- Сфотографироваться так, чтобы было видно лицо и оригинально оформленная подпись.
- Переслать готовый снимок – сигну адресату.
На бумаге
Создать сигну можно на обычном листе бумаги. Так часто поступают знаменитости или обычные пользователи всемирной паутины, чтобы подтвердить достоверность свой электронной страницы. Такие сигны появились одними из первых, поэтому при их создании не возникает никаких проблем. Итак:
- Написать на листке бумаги свое имя, имя парня или электронный адрес разборчивым почерком.

- Сфотографировать себя с оформленным листком бумаги так, чтобы отчетливо виднелось лицо.
- Переслать сигну адресату.
Без лица
Такие реальные фото добавляют интригу, а чаще используются на сайтах о похудении, внешнем перевоплощении и т.д. Для создания сигна без лица тоже можно использовать лист бумаги с оформленной тематической надписью. Вот инструкция:
- Оформить надпись на бумаге.
- Закрыть лицо, выполнить фото.
- Переслать сигну адресату.
Фото с сигной
Видео
Что такое сигна в ВК (Вконтакте)?
Сигна в ВК – это просто фотография человека с листком бумаги, на котором обычно написано чье-то имя. Часто надписи делают на ладони и груди. Это вроде как должно придавать большую ценность написанному.
Само слово произошло от английского “Signature”, что означает “Подпись“.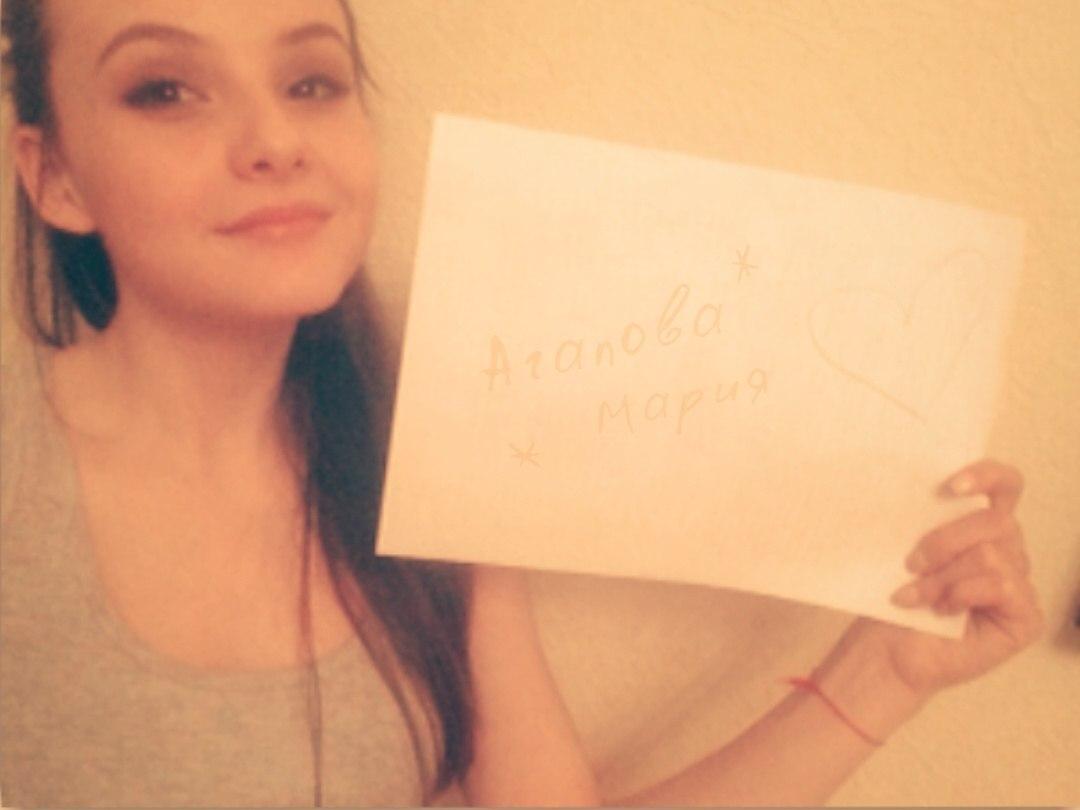 Когда вас просят сделать сигну – это значит, что вы должны сфотографироваться с именем этого человека.
Когда вас просят сделать сигну – это значит, что вы должны сфотографироваться с именем этого человека.
Правильная сигна – это та, которая написана от руки.
Фото
Это классический способ. Просто сделайте фото с именем, к примеру, на табличке.
Видео
Это когда записывают на видео целое поздравление или обращение, и держат в руках сигну с именем человека, которому это адресовано.
Такую сигну можно сделать за деньги. Существуют сайты, на которых за пару баксов люди готовы сделать надпись с вашим текстом на своем теле. Для примера я заказал у JungleBoyz надпись в виде адреса моего сайта.
Вот такая видео-сигна получилась:
Как сделать сигну в ВК? Используйте ботов Вконтакте!
Вконтакте есть специальные боты, которые делают надписи в автоматическом режиме. В качестве “холста” выступают фотографии знаменитостей.
Примеры ботов:
- Botsigna
- Signabot
- Signaphoto
Вот такие ВК сигны у них получаются:
Что такое сигна фото | NUR.
 KZ Сигна: Avatan Plus
KZ Сигна: Avatan PlusСигна — способ идентификация и подтверждения реальности собеседника в социальных сетях. Для этого человек фотографируется с надписью на листочке, говорящей о подлинности аккаунта. Узнайте, откуда взялось это понятие, для чего нужно и как сделать собственную сигну.
Что такое сигна
Слово «сигна» происходит от английского sign (‘знак’).
Изначально сигны появились в соцсетях для подтверждения реальности собеседника. Чтобы удостовериться, что это не фейковая страница, просили сфотографироваться с надписью на листочке, которая подтвердит подлинность аккаунта.
К такой идентификация прибегают знаменитости, чтобы отделить настоящий аккаунт от сотен поддельных.
Помимо этого, сигны используют для подтверждения авторства различных творческих работ, в частности фотографий. Водяные знаки или подпись можно затереть в фотошопе, а вот убрать сигну со снимка совсем не просто.
Читайте также
Как использовать Snapchat на Android: возможности и эффекты
Некоторые начинающие блогеры, мечтающие о раскрутке в социальных сетях, скрывают свое лицо за сигной, создавая атмосферу таинственности и тем самым подогревая к себе интерес.
Со временем появилось и новое применение для личной подписи. Для многих людей сигна — это способ выразить свои чувства и поделиться сокровенным.
Как сделать сигны
Перед тем как сделать сигну, определитесь, какая подпись и для чего вам нужна.
Для подтверждения личности или отправки личного послания подойдет сигна на бумаге или на теле.
Сигна: Сигны — BeOnПодключите фантазию — даже простая сигна с именем станет произведением искусства, если ее правильно оформить. Экспериментируйте со шрифтами и рисунками.
Учитывайте основные правила нанесения сигны. Она должна быть:
Читайте также
Кем могут представляться финансовые мошенники в Казахстане
- разборчивой;
- выделяться на фоне тела или бумаги;
- оформленной зеркально, чтобы читаться на снимке.
Не обязательно фотографироваться с лицом, если только это не было основным условием предоставления сигны. Скройте лицо за надписью или вовсе оставьте за кадром.
Сигнами можно подтверждать не только фото, но и видео. Во время съемки вы можете показать на камеру знак на теле или бумаге.
Во время съемки вы можете показать на камеру знак на теле или бумаге.
Для проверки подлинности фото с сигной используют специальные программы. Также пригодится наблюдательность — некоторые подделки можно заметить невооруженным глазом.
Защитите свое творчество и свой аккаунт в социальных сетях фото с сигной. Это интересное новшество станет оригинальным способом общения с друзьями и выделит вас среди других.
Читайте также
Как удалить аккаунт в Facebook навсегда и деактивировать его
Оригинал статьи: https://www.nur.kz/leisure/interesting-facts/1710488-cto-takoe-signa-foto/
Плесень и покровные дефекты на сигарах
Мы привыкли бороться с плесенью дома, где бы она ни появилась. Но вряд ли нам удастся избежать ее повсюду. Как распознать плесень на сигарах и как на нее правильно реагировать, обсудим в этой статье.
Прежде всего, нужно различать разные дефекты на покровном листе, а также отличать вредную плесень от такого явления как “шлейф” (плюм, англ.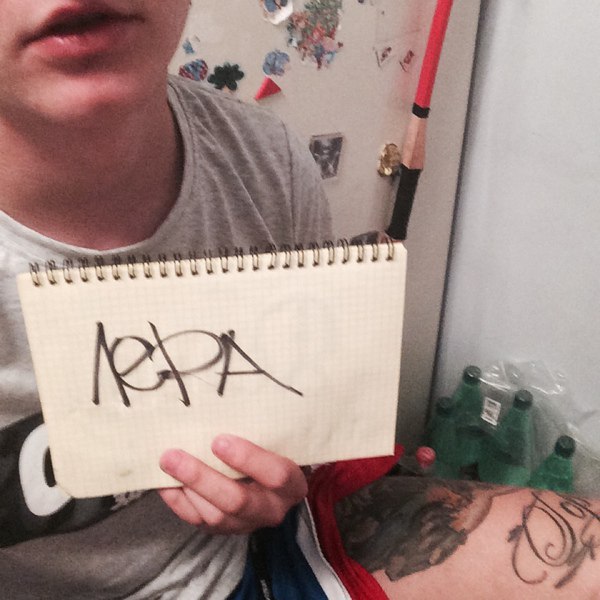
На самом деле «шлейф» — это не что иное, как кристаллизация солей в составе масел табачного листа. Некоторые наблюдатели отмечают, что кристаллики искрятся в луче света, направленном под углом 90 градусов.
Плюм выглядит как белые точки или тонкий белый слой
Вы можете встретить и другие дефекты покровного листа, которые слегка портят эстетическое впечатление от витолы, но чаще всего никак не влияют на её вкусовые качества. Так, иногда на сигаре можно заметить небольшие 
Зеленые участки на сигарном покрове не влияют на ее качество
Другим безобидным недостатком являются пятнышки от воды. Они обычно представляют собой желтовато-белые круги на фоне ровного коричневого покрова. Хотя ведутся споры о том, почему появляются эти пятна, одно остается бесспорным — они абсолютно безопасны.
Круглые гладкие пятна, высветляющие покров, — пятна от воды
Что касается плесени, то здесь ситуация может оказаться довольно опасной. Прежде всего, плесень может быть различных цветов, таких как зеленый, синий, белый и слегка желтоватый. Во-вторых, она выглядит «мохнатой» и растет гораздо быстрее, чем образуется плюм.
Если Вы попытаетесь стереть с сигары плесень, она оставит следы. Если так, то не ограничивайтесь одним вытиранием. Капните на чистую ткань пару капель спирта и протрите еще раз. Такие сигары следует выкурить как можно скорее, иначе плесень быстро распространится, и, возможно, сигару придется выбрасывать.
Наиболее распространенным видом является белая плесень, которую иногда путают с плюмом, по сравнению с которым она выглядит более «грязным» и объемным пятном.
Плесень более объемная и мохнатая
Нужно помнить, что плесень растет не только снаружи, но и пускает “корни” внутрь, таким образом разрушая сигару. Если сигара сильно заплесневела, ее рекомендуется выбросить.
Необходимо обработать спиртом хьюмидор, в котором она лежала. Лучше сделать это дважды, а затем проветрить хьюмидор в течение суток. Не забудьте наполнить увлажнитель дистиллированной водой, не используйте водопроводную.
Почему появляется плесень и как ее предотвратить? Очевидно, что плесень появляется из-за повышенной влажности и/или температуры в Вашем хьюмидоре. Контролируйте ее как можно чаще. Если влажность будет больше 80%, плесень появится наверняка. Температура хранения сигар не должна превышать 25 градусов. Достаточная циркуляция воздуха в хьюмидоре также необходима, чтобы избежать переувлажнения сигар.
Цифровой гигрометр в хьюмидоре — не роскошь, а средство предотвращения плесени
Таким образом, плесень на сигарах лучше стараться предотвратить, поддерживая оптимальную влажность (72%) и температуру (не более 20 С) в хьюмидоре. Если же Вы заметили ее, нужно вовремя принять меры. Вам решать, избавляться ли от зараженных сигар, или оставить их. Наверное, лучшим решением будет поскорее выкурить их.
Преобразование сигналапри старении листьев: прогресс и перспектива
растений (Базель). 2019 Октябрь; 8 (10): 405.
Салман Ахмад
1 Научно-исследовательский институт табака Китайской академии сельскохозяйственных наук, Циндао 266101, Китай; moc.oohay@aftifas
2 Отдел селекции и генетики растений, Ядерный институт продовольствия и сельского хозяйства, Тарнаб, Пешавар П.О. Box 446, Пакистан
Yongfeng Guo
1 Институт исследования табака Китайской академии сельскохозяйственных наук, Циндао 266101, Китай; мок.oohay @ aftifas
1 Научно-исследовательский институт табака Китайской академии сельскохозяйственных наук, Циндао 266101, Китай; moc.oohay@aftifas2 Отдел селекции и генетики растений, Ядерный институт продовольствия и сельского хозяйства, Тарнаб, Пешавар П.О. Box 446, Пакистан
Поступила 14 сентября 2019 г .; Принято 8 октября 2019 г.
Лицензиат MDPI, Базель, Швейцария. Эта статья представляет собой статью в открытом доступе, распространяемую в соответствии с условиями лицензии Creative Commons Attribution (CC BY) (http: // creativecommons.org / licenses / by / 4.0 /). Эту статью цитировали в других статьях в PMC.Abstract
Старение листа — это дегенеративный процесс, который контролируется генетически и включает ремобилизацию питательных веществ до отмирания тканей листа. Возраст является ключевым фактором, определяющим этот процесс, наряду с другими факторами, вызывающими старение. На клеточном уровне различные гормоны, сигнальные молекулы и факторы транскрипции участвуют в регуляции старения. Этот обзор суммирует недавний прогресс в понимании сложности процесса старения с уделением основного внимания восприятию и передаче сигналов старения, а также последующих регуляторных событий.Будут обсуждены будущие направления в этой области и потенциальные применения связанных методов в улучшении сельскохозяйственных культур.
Ключевые слова: старение листа, возраст, передача сигнала, рецептор-подобная киназа, LRR-RLK, каскад киназ MAP, фактор транскрипции
1. Введение
Старение листа — это совокупный ответ множества факторов, действующих на последних стадиях. жизненного цикла растений. Начало старения приписывается потере хлорофилла в клетке и впоследствии распространяется на распад макромолекул, включая белки, углеводы, липиды и нуклеиновые кислоты [1,2,3].В то же время вырожденные молекулы перерабатываются и перераспределяются в запасающие ткани и органы в зависимости от жизненного цикла растений. Растения с коротким жизненным циклом восстанавливают перемещенные питательные вещества в свои семена и плоды, в то время как у многолетних растений, таких как деревья, стебель и корень служат органами хранения, которые впоследствии используются для появления новых листьев и цветов в следующем сезоне [4,5 , 6]. Постепенная деградация хлоропластов и потеря фотосинтетической активности оставляют желтый след на поверхности листа от кончика до основания.Это преимущественное разложение хлорофилла приводит к изменению цвета монокарпических растений с зеленого на золотисто-желтый по мере созревания зерна перед сбором урожая. Кроме того, распад хлорофилла в сочетании с синтезом новых пигментов, таких как антоцианы и фенолы, из-за гидролиза макромолекул дает заметный диапазон цветов осенних листьев лиственных деревьев [7].
Старение листьев можно рассматривать как процесс дегенерации и гибели клеток, но в глубине души это систематический механизм, который работает под жестким генетическим контролем, чтобы гарантировать выживание растения и накопление питательных веществ для следующего поколения.Инициирование старения, генетическое разнообразие симптомов старения и перемещение питательных веществ во времени, пространстве и количественно — это проявления генетически контролируемой программы. На клеточном и субклеточном уровнях различные гормоны, рецепторы и факторы транскрипции вносят свой вклад в регуляцию и выполнение старения. Исследования на генетическом и молекулярном уровнях демонстрируют широкий спектр сложного и высокоорганизованного синдрома старения [8,9]. Генетические подходы, сфокусированные на мутантах с измененной фенотипической экспрессией листа во время старения, предполагают значительную роль регуляторных элементов, как положительных, так и отрицательных, для включения и выключения прогрессирования старения [10,11].Изучение и характеристика межклеточных и внутриклеточных сигнальных путей, регуляторных факторов и профилей дифференциальной экспрессии генов дает исчерпывающее молекулярное понимание синдрома старения. Передача сигналов является очень важным аспектом старения листа, так как инициация или индукция процесса напрямую зависит от различных внешних или внутренних сигналов / сигналов. Засуха, засоление, высокая / низкая температура, дефицит питательных веществ, низкая интенсивность света и инфекция патогенными микроорганизмами — это различные типы сигналов окружающей среды, которые подталкивают растение к старению.Помимо внешних факторов, эндогенные гормоны также участвуют в сигнальных каскадах старения. Запущенные пути в результате действия внешних и внутренних сигнальных факторов формируют сложную сеть регуляции старения листа [8,9].
Здесь, в этом обзоре, мы суммируем все те существенные факторы и компоненты, которые участвуют в передаче сигналов, с учетом современных исследований и достижений, сделанных в этой важной фазе развития старения.
2.Гормоны как регулирующие старение сигналы
Как только клетка воспринимает сигналы / сигналы старения, запускается серия событий передачи сигнала с участием различных сигнальных компонентов на разных уровнях регуляции, и между различными сигнальными компонентами происходят значительные взаимодействия [8, 9]. Гормоны играют важную роль в регуляции старения листьев как в зависимости от возраста, так и в результате стресса. Сообщалось, что почти все основные фитогормоны действуют в сигнальных путях старения.Этилен, абсцизовая кислота, брассиностероиды, жасмоновая кислота, салициловая кислота, цитокинины, ауксин и гибберелловая кислота, очевидно, контролируют старение листьев в различных исследованиях [12,13,14].
Этилен играет множество ролей в различных процессах развития, таких как пролиферация, удлинение, размер клеток, опадение и созревание плодов, старение и стрессовые реакции на абиотические / биотические факторы [12]. Этилен играет роль в передаче сигналов, связанных со старением, и известен как гормон, способствующий старению [15].Этиленовый сигнальный каскад, как было предсказано, формирует прямой путь, включающий мембраносвязанные рецепторы на одном конце, за которыми следуют ряд положительных / отрицательных регуляторов и нижестоящие факторы транскрипции на другом конце пути [16]. Применение этилена ускоряет старение листьев и цветов, ингибируя синтез этилена, что приводит к задержке старения [17]. Исследования экспрессии генов выявили повышенное количество транскриптов, связанных с синтезом этилена и передачей сигналов во время старения листьев в зависимости от возраста [16].Мутантные нечувствительные к этилену2 ( ein2 ) растения обнаруживают замедленное старение листьев из-за низкой экспрессии 21 гена, связанного со старением ( SAGs ), которые кодируют ферменты полигалактуроназы и пектинэстеразы, ответственные за разложение клеточной стенки [18]. EIN3, сигнальная молекула, расположенная ниже EIN2, как было показано, положительно регулирует старение листа посредством активации двух факторов транскрипции, способствующих старению, ORE1 и AtNAP [19]. Было подтверждено, что этиленовый сигнальный каскад, опосредованный старением, функционирует в зависимости от возраста, о чем свидетельствует обработка экзогенным этиленом на зрелых и молодых листьях, что приводит к раннему и нормальному старению соответствующих листьев [12, 20].
Абсцизовая кислота (АБК) является важным фитогормоном, регулирующим широкий спектр процессов роста и развития, включая прорастание, покой, зрелость, эмбриогенез, закрытие устьиц, регуляцию роста побегов и корней, созревание плодов, старение листьев, опадение и стрессовые реакции, которые включают засуху, засоление и низкую температуру [12,21]. Механизмы этих функций контролируются различными сигнальными механизмами, например, путями Ca 2+ , ROS, G-Protein, SnRK2, PP2C и MAPK [14,22,23].Ряд исследований АБК ясно демонстрируют ее роль как положительного регулятора старения листьев. Исследования на микрочипах выявили повышенную экспрессию генов, участвующих в биосинтезе и сигнальных ответах ABA во время старения у Arabidopsis [18]. Исследования также сообщили о повышенном уровне эндогенной АБК и активации генов, связанных с передачей сигналов АБК во время старения у различных растений, таких как кукуруза, табак, рис, овес и арабидопсис [11,24]. Было показано, что рецептор ABA PYL9 способствует устойчивости к засухе и старению листьев у Arabidopsis [25].В условиях обезвоживания АБК контролирует потерю воды за счет закрытия устьиц и предотвращает раннее старение растения [26]. Связанный со старением ген 113 ( SAG113 ), кодирующий протеинфосфатазу 2C (PP2C), действует как негативный регулятор передачи сигналов ABA, подавляя закрытие устьиц, что приводит к усыханию листьев [12, 24, 27].
Брассиностероиды (BR) выполняют многофункциональные функции в растениях, такие как прорастание, цветение, опадение, старение и в ответ на различные стрессы [12,28,29].Путь передачи сигналов BR состоит из трех гомологов BRI1-ассоциированной рецепторной киназы 1 (BAK1), действующих как корецепторы мембраносвязанной рецепторной киназы 1 (BRI1) для восприятия лигандов BR [28]. Роль брассиностероидов в положительной регуляции старения листьев была отмечена ускорением старения посредством внешней обработки и задержкой старения у мутантных растений BR [12,30]. Применение эпибрассинолида (eBL) на отделенных листьях пшеницы показало замедленное и ускоренное старение листьев, обеспечиваемое в низкой и высокой концентрации, соответственно [30].Экспрессия генов, кодирующих сигнальные белки BR BRI1 киназы 1 (BKI1) и BZR1 в стареющих листьях, была значительно подавлена в мутанте раннего старения риса ospls1 [31]. Когда отделенные листья Pisum sativum обрабатывали 24-эпибрассинолидом (EBR), старение усиливалось, а содержание свободных жирных кислот увеличивалось, что позволяет предположить, что BR могут регулировать старение листьев посредством изменения липидного состава клеток [32].
Жасмоновая кислота (JA) и метилжасмонат (MeJA) участвуют в различных процессах роста и развития растений, включая старение и различные реакции на биотический и абиотический стресс [12,33].Участие жасмоната в регуляции старения было впервые обнаружено, когда обработанные MeJA отдельные листья овса показали ускоренное старение листьев [34]. У Arabidopsis 14 из 125 линий-ловушек-энхансеров, ассоциированных со старением, обнаружена JA-индуцированная экспрессия репортерного гена [35]. В стареющих листьях продуцируется в четыре раза больше эндогенного JA по сравнению с не стареющими, а обработка JA инициировала старение прикрепленных и отслоившихся листьев [36]. Сообщалось о повышенной экспрессии генов синтеза JA ( LOX3 , OPR3 , AOC1 и AOC4 ) и сигнальных генов ( MYC2 , JAZ1 , JAZ6 и JAZ8 ). -зависимое старение листьев [16,37].Снижение экспрессии генов, связанных с фотосинтезом, происходило после экзогенного применения JA и MeJA [12]. Применение MeJA также увеличивало экспрессию генов, связанных с возрастным старением, включая SEN4 , ERD1 и SAG21 [12,38,39]. Фактор этиленового ответа, BrERF72, предположительно участвует в регуляции JA-опосредованной активации экспрессии BrLOX4 , BrAOC3 и BrOPR3 во время JA-промотируемого старения листьев в цветущей капусте китайской [40].
Салициловая кислота (SA) действует как сигнальная молекула для регулирования процессов развития, включая прорастание семян, цветение, старение и ответы на абиотические / биотические стрессы [41]. Предполагается, что как положительный регулятор онтогенетического старения листа, SA участвует как в инициации, так и в прогрессировании старения листа [16,42]. Анализ транскриптома и микроматрицы сообщил о том, что ряд генов биосинтеза SA, передачи сигналов и чувствительных генов активируются во время старения листьев в процессе развития [16].Исследование профилей экспрессии генов Arabidopsis дикого типа и мутанта NahG (с пониженной SA) показало зависимость многих SAG от сигнальных путей SA [16,18]. Более недавнее исследование показало, что фактор транскрипции WRKY75 положительно регулирует старение листьев, способствуя продукции SA и подавляя удаление H 2 O 2 , частично путем подавления транскрипции CATALASE2 [43].
Цитокинины (ЦК) — важные фитогормоны, играющие важную роль в процессе роста и развития растений.ЦК также известны как гормоны, замедляющие старение, ингибирующие деградацию хлоропластов при экзогенном или эндогенном применении [11,44]. Анализ транскриптома показал, что во время старения снижается накопление в транскриптах генов синтеза ЦК ( изопентилфосфотрансфераза ( IPT ) и цитокининсинтаза ) и повышается уровень генов, кодирующих ферменты разрушения цитокининов, такие как цитокининоксидаза и цитокинин, инактивирующий N- и O-гликозилазы, присутствовали [18].Когда ген синтеза CK IPT экспрессируется под контролем специфичного для старения промотора SAG12, значительная задержка старения листьев наблюдается у большого числа видов растений [44,45]. Из-за точно настроенной специфической для старения экспрессии IPT в этих исследованиях, индуцированное старением повышение уровней эндогенных ЦК подавляло или обращало старение листьев однажды инициированное [45]. Помимо естественного старения, повышенные уровни ЦК в растениях, экспрессирующих SAG12-IPT , часто вызывали повышенную устойчивость к стрессам и замедленное старение, вызванное стрессовыми условиями [45].Подавление вызванного засухой старения листьев наблюдали у ползучей полевицы с повышенной экспрессией IPT , что вызвало активацию антиоксидантных ферментов, включая пероксидазу, супероксиддисмутазу и каталазу [12,46]. Поддержание фотосинтетической активности и увеличение роста корней при стрессе влаги наблюдалось у растений, сверхэкспрессирующих гены синтеза цитокининов [46,47]. Было показано, что у Arabidopsis сигнальный путь CK включает His протеинкиназы (AHK) в качестве рецептора CK, носителя фосфора B-типа и регулятора ядерного ответа A-типа (ARR), которые действуют ниже рецепторов, контролируя старение листьев. [16,48].Мутация с усилением функции в рецепторе CK AHK3 вызвала задержку старения листьев, в то время как мутация с потерей функции в этом белке привела к снижению чувствительности к цитокинин-зависимой задержке старения листьев [49]. Кроме того, 71 активированный и 11 отрицательно регулируемых генов, непосредственно чувствительных к ЦК у Arabidopsis, о которых сообщается в профиле экспрессии по всему геному, могут потенциально участвовать в передаче сигналов старения, опосредованной ЦК [50].
Ауксины играют важную роль в росте и развитии растений и, как предполагается, действуют как негативный регулятор старения листьев [51].Обработка отделившихся листьев Arabidopsis ауксином вызвала снижение экспрессии SAG12 , гена-маркера онтогенетического старения [52]. Повышенное накопление эндогенного ауксина в растениях Arabidopsis, сверхэкспрессирующих ген биосинтеза ауксина YUCCA6 , показало задержку старения листьев и снижение экспрессии SAGs [53]. Другое исследование показало, что, когда ген SAUR39 малой РНК-ауксина был сверхэкспрессирован в рисе, снижение транспорта ауксина и доступность свободной IAA наблюдались в трансгенных растениях, которые проявляли фенотипы раннего старения [54].Более того, было идентифицировано, что репрессоры генов ответа на ауксин, AUXIN RESPONSE FACTORs 1 и 2, участвуют в регуляции старения и опадания листьев Arabidopsis [55]. Нокаут-мутант Arabidopsis ORESARA14 , кодирующий ARF2, обнаруживал задержку старения листьев [56]. Роль ауксина в старении, однако, кажется более сложной, поскольку сообщалось о повышенном уровне ауксина (ИУК) в листьях старения [57], а индуцированное ИУК производство этилена в листьях табака способно противостоять старению. задерживающая активность ИУК [58].
Гибберелловая кислота (GA) способствует удлинению стеблей и листьев, прорастанию семян, цветению, образованию плодов и семян и реагированию на различные стрессы [59,60]. Сообщается, что GA является фитогормоном, замедляющим старение, поскольку уровень его активной формы снижается по мере прогрессирования старения в процессе развития [13]. Роль GA в замедлении старения была впервые показана при обработке отслоившихся тканей листа Taraxacum officinale GA [61]. Ряд исследований указывает на наличие свободного GA (GA4 и GA7), ингибирующего старение листьев; хотя одно исследование показало, что задержка старения листьев у Paris polyphylla , вызванная GA, была вызвана провоцирующим действием АБК [62,63].Экспрессия GA-индуцибельного AtGA2OX2, который кодирует GA 2-оксидазу2, которая деактивирует GA, увеличивается в 18 раз во время старения [16]. Мутанты Arabidopsis, дефицитные по биосинтезу GA или передаче сигналов GA, проявляют фенотипы замедленного старения, подтверждая роль GA как отрицательного игрока в регуляции старения листьев.
3. Факторы транскрипции в пути передачи сигнала старения
Восприятие и передача сигналов / сигналов, вызывающих старение, приводят к резким изменениям в экспрессии генов, которые управляют реализацией синдрома старения и в конечном итоге приводят к гибели клеток [37, 64,65].Гены, которые активируются во время старения, обычно называются генами, связанными со старением ( SAG, ), тогда как гены с подавленным паттерном экспрессии, такие как гены фотосинтеза, называются генами, подавляющими старение ( SDG, ). Было показано, что факторы транскрипции (TF) как регуляторные белки, служащие переключателями в процессе дифференциальной экспрессии генов, участвуют как в стимулировании, так и в ингибировании старения растений [9,66,67]. Сообщается, что ряд семейств TF являются регуляторами старения листьев [18,27,68].На основании ДНК-связывающих доменов среди 2403 генов, кодирующих TF в геноме Arabidopsis, было показано, что по меньшей мере 287 TF в 34 семействах генов связаны со старением, включая членов NAC, WRKY, MYB, C2h3, APE2, bZIP и семейства генов HB [27,68,69,70].
NAC представляет собой большое семейство TF и широко ассоциировано с регуляцией процесса старения листьев [19,71,72,73]. Некоторые гены NAC TF, такие как AtNAP, ORE1, ORS1, ANAC016 и ATAF1, являются положительными регуляторами старения, и их избыточная экспрессия обычно приводит к преждевременному старению, в то время как подавление их экспрессии вызывает замедленное старение у Arabidopsis [64,74,75,76].Напротив, негативные регуляторы старения в семействе NAC, которые дают фенотип замедленного старения при сверхэкспрессии, также описаны, такие как VNI2 (VND-INTERACTING2) и JUB1 (JUNGBRUNNEN1) [77,78,79]. Во время старения NAC TF обычно образуют сложные регуляторные сети, контролируя другие NAC и взаимодействуя с NAC или другими TF, чтобы регулировать экспрессию генов-мишеней [73,80].
Другим важным семейством TF, регулирующим старение листа, является WRKY, который участвует во множестве процессов роста и развития, включая старение и реакции на стресс.Роли WRKY6, WRKY18 и WRKY22 / WRKY29 описаны в защитных реакциях и регуляции старения [68,81,82,83]. WRKY53 — фактор транскрипции, способствующий старению, который управляет экспрессией ряда SAG , некоторые из которых связаны со стрессовыми ответами и атаками патогенов [84,85]. WRKY22 был идентифицирован как мишень для фактора транскрипции WRKY53, который играет положительную роль в регуляции индуцированного темнотой старения, тогда как его нокаут-мутант проявляет замедленное старение листьев [85,86].Кроме того, было подтверждено, что WRKY54 и WRKY70 играют негативную роль в регуляции старения [42,85].
Члены из других семейств факторов транскрипции, которые регулируют старение, включают позитивные регуляторы AP2, AUXIN RESPONSE FACTOR ARF2, фактор связывания C-повтора CBF2, фактор связывания G-Box GBF1, MYB-подобный фактор транскрипции AtMYBL [56,64,87,88] и негативные регуляторы, такие как регулятор ответа цитокининов B-типа ARR2 [49,64].
4. Восприятие и передача сигналов, связанных со старением
В течение последних двух десятилетий процесс старения листьев широко изучался на модельном растении Arabidopsis с большим упором на роль различных гормонов и идентификацию регуляторных генов. особенно те, которые кодируют факторы транскрипции [8,9].Однако молекулярные механизмы того, как сигналы / сигналы старения воспринимаются / воспринимаются и передаются в клетку и в конечном итоге приводят к переключению изменений экспрессии генов и запускают выполнение старения листа, все еще неясны. В дополнение к возрасту, который считается основным сигналом старения растений, ряд стимулов окружающей среды также играют роль в запуске старения. К ним относятся засуха, дефицит питательных веществ, темнота, экстремальные температуры, инфекция патогенными микроорганизмами и гормоны растений, включая этилен, абсцизовую кислоту, жасмоновую кислоту и салициловую кислоту [44,89,90].Восприятие и передача различных реплик / сигналов в конечном итоге приводит к подобному «синдрому старения» [64], и ожидается значительное перекрытие и перекрестные переговоры между этими сигнальными путями.
Многие из этих сигналов / сигналов воспринимаются / воспринимаются рецепторными белками на поверхности клетки и затем передаются через плазматическую мембрану, чтобы активировать передачу сигнала внутри клетки, чтобы активировать генную регуляторную сеть старения. Информация о возрастных и опосредованных окружающей средой сигнальных путях старения ограничена и не может раскрыть сложную регуляторную сеть этого процесса.Однако большое количество генов, потенциально участвующих в передаче сигналов, было идентифицировано с помощью транскриптомного анализа. Основные группы этих генов включают митоген-активируемую протеинкиназу (MAPK) и рецептор-подобные киназы (RLK) [37, 65, 68].
4.1. Рецептороподобные киназы (RLK) в Leaf Senescence
Рецептороподобные киназы растений (RLK) представляют собой рецепторы клеточной поверхности, которые обладают уникальными структурными особенностями [91,92]. RLK обычно имеет N-концевой внеклеточный связывающий домен для связывания лиганда, трансмембранный домен, который охватывает плазматическую мембрану, и цитоплазматический киназный домен, который обычно функционирует в передаче сигнала через фосфорилирование нижестоящих компонентов для активации регуляторной сети [91].Внеклеточный домен участвует в восприятии лигандов [68,93]. Впоследствии, после связывания лигандов, функциональная особенность RLK заключается в аутофосфорилировании внутриклеточного компонента, который активирует нижестоящую регуляторную сеть и изменения экспрессии генов [82,93,94]. С более чем 600 членами, кодируемыми геномом Arabidopsis, RLK представляют собой крупнейшее суперсемейство белков в растениях с широко распространенной функциональной ролью в развитии, росте, устойчивости к патогенам, самонесовместимости и гормональных ответах [95].Было охарактеризовано более 20 различных типов внеклеточных доменов RLK, включая богатые лейцином повторы (LRR), повторы эпидермального фактора роста, самонесовместимость (S) и лектиновые домены [93,96,97]. Крупнейшее подсемейство RLK у Arabidopsis — это протеинкиназа, подобная рецептору с богатыми лейцином повторами (LRR-RLK), насчитывающая более 200 членов [95]. Внеклеточный связывающий домен LRR-RLKs содержит различное количество богатых лейцином повторяющихся единиц, которые обычно имеют длину 24 аминокислоты [91].Хотя большое количество белков LRR-RLK было идентифицировано у различных видов растений, включая арабидопсис, томат, рис, картофель и тополь [95,98,99,100], до сих пор охарактеризовано лишь ограниченное количество LRR-RLK. играть роль в развитии растений и реакции на стресс [91].
RLK функционируют, воспринимая сигналы на поверхности клетки и передавая сигналы через плазматическую мембрану, чтобы активировать передачу сигнала внутри клетки, что делает их хорошими кандидатами в рецепторы сигналов, вызывающих старение.Фактически, ряд белков семейства LRR-RLK был охарактеризован как регулятор старения листьев. Используя подход дифференциального отображения мРНК, ген LRR-RLK, названный связанной со старением рецептор-подобной киназой (SARK), был выделен из фасоли ( Phaseolus vulgaris cv Bulgarian), чтобы иметь повышенные уровни как транскрипта, так и накопления белка во время старения листа. [101]. Промотор P SARK был использован для управления экспрессией IPT и для повышения устойчивости растений к стрессу [102].Однако о функции SARK в регуляции старения не сообщалось. Другой ген LRR-RLK с повышенной регуляцией старения, названный GmSARK, был позже выделен в сое ( Glycine max ) и, как было обнаружено, участвует в регуляции старения листьев [103]. Было показано, что GmSARK сои и его гомолог у Arabidopsis, AtSARK, действуют как положительный регулятор старения листьев. Индуцируемая сверхэкспрессия GmSARK или AtSARK вызывала преждевременное старение, тогда как старение листьев задерживалось у растений со сниженной экспрессией этих генов [85,104].Совсем недавно было показано, что белок LRR-RLK из мха ( Physcomitrella patens ), гомологичный SARK фасоли, названный PpSARK, действует как негативный регулятор старения мха [105]. Следует отметить, что хотя все вышеупомянутые SARK , как сообщалось, имеют повышенную регуляцию старения, только PpSARK был выделен на основании сходства последовательностей, и он имеет относительно высокую гомологию с SARK фасоли [104,105].
Помимо SARK, ряд других белков LRR-RLK был охарактеризован на предмет их роли в старении листьев у Arabidopsis.АБК-воспринимающая мембраносвязанная рецепторная киназа RPK1 (РЕЦЕПТОРНАЯ ПРОТЕИНКИНАЗА 1) была идентифицирована как обладающая положительной регуляторной функцией при возрастном и АБК-опосредованном старении листьев [85,106]. Нокаут-мутанты RPK1 обнаруживают значительную задержку как возрастного, так и индуцированного АБК старения [106]. Совсем недавно было обнаружено, что член из подсемейства LRR-RLK киназ, подобных рецепторам соматического эмбриогенеза, SERK4 действует как негативный регулятор в путях передачи сигналов старения листьев [107]. Семейство SERK арабидопсиса состоит из пяти членов (SERK1-5), и белки SERK, как сообщается, действуют как корецепторы в различных сигнальных путях, регулируя различные процессы развития и реакции на стресс [108].Интересно, что AtSARK был также идентифицирован как киназа 3 нечувствительного рецептора CLAVATA3 (CIK3) вместе с LRR II-RLK CIK1, CIK2 и CIK4 в той же подгруппе, чтобы функционировать как корецепторы в пути CLAVATA для регуляции гомеостаза стволовых клеток у арабидопсиса. [109]. Было показано, что один корецептор может работать с несколькими рецепторами в разных сигнальных путях [108, 110]. AtSARK и SERK4, вероятно, являются частью рецепторных комплексов, функционирующих вместе с другими LRR-RLK в регуляции старения.
LRR-RLK, индуцированная старением, рецептор-подобная киназа (SIRK) была охарактеризована, чтобы показать характер экспрессии, специфичный для старения листа [82]. Выявлено, что более 40 генов рецептор-подобных киназ связаны со старением листьев арабидопсиса [68]. Недавнее исследование выявило киназу рецептора риса OSBBS1 / OsRLCK109, которая принадлежит к подсемейству RLCK и не имеет внеклеточного домена, который играет важную регуляторную роль в старении листьев и ответах на солевой стресс. Мутант bbs1 проявлял гиперчувствительность к солевому стрессу и фенотипам раннего старения листьев [111].Роли большего количества LRR-RLK и других типов RLK еще предстоит выяснить.
4.2. Активированная митогеном протеинкиназа в передаче сигналов старения
Передача сигнала, инициированная RLK, часто осуществляется внутри клетки посредством фосфореле. Каскад митоген-активируемой протеинкиназы (MAPK) (MAPKKK-MAPKK-MAPK) является одним из наиболее важных путей передачи сигнала у растений. Участие каскадных путей MAPK в широком диапазоне клеточных процессов, таких как деление клеток, дифференцировка, ответы на биотические / абиотические стрессы и гормоны, было показано ранее [112].Более раннее исследование идентифицировало 9 генов MAPKKK, 3 MAPKK и 3 MAPK как связанных со старением листьев Arabidopsis [68]. У кукурузы ZmMPK5 было идентифицировано как связанное со старением листьев и восстановлением после низкотемпературного стресса [113].
У Arabidopsis, MAPKKK18, как сообщается, является положительным регулятором старения листьев, а регуляторная роль MAPKKK18 в старении зависит от его киназной активности и передачи сигналов ABA [14]. Точно так же у риса SPOTTED LEAF3 (SPL3), также известный как OsMAPKKK1, положительно регулирует старение листьев посредством пути передачи сигналов ABA [114].Было показано, что для MAPKK MAPKK9 (MKK9) играет важную регулирующую роль в старении листьев Arabidopsis. Когда MKK9 , но не неактивная форма киназы MKK9KR , сверхэкспрессировалась, трансгенные растения проявляли фенотип преждевременного старения листьев. С другой стороны, старение отделившихся листьев растений с нулевым мутантом mkk9 и было задержано [90]. Другой MAPKK у Arabidopsis, EDR1, был идентифицирован как негативный регулятор защитных реакций и этилен-индуцированного старения листьев [115, 116].MAPKK кукурузы, ZmMEK1, также был охарактеризован как негативный регулятор старения. Накопление транскриптов ZmMEK1 и индуцировалось во время индуцированного темнотой старения листьев кукурузы, а экспрессия доминантно-отрицательного мутанта ZmMEK1 в Arabidopsis индуцировала SA-зависимое старение листьев [117]. Сигналы обычно далее трансдуцируются в MAPK посредством фосфорилирования до достижения факторов транскрипции, которые изменяют экспрессию генов. У Arabidopsis MAPK6 (MPK6) был идентифицирован как цель MKK9 в каскаде MAPK.Когда и MKK9 , и MPK6 экспрессировались в протопластах нулевого мутанта mkk9 , было показано, что MKK9 способен фосфорилировать MPK6 у Arabidopsis. Киназная активность усиливалась присутствием конститутивно активной формы MPKK9 (MKK9EE). И что более важно, mpk6 нулевые мутанты фенокопировали mkk9 с задержкой старения листьев, и роль MKK9 в стимуляции старения частично зависела от MPK6 [90]. У кукурузы MAPK ZmSIMK1 был идентифицирован как прямая мишень для MAPKK ZmMEK1, и было показано, что каскад ZmMEK1-ZmSIMK1 и его модуляция уровней SA играют важную роль в регуляции старения листьев [117].
Сигнальный каскад киназы MAP существенно изменяет экспрессию генов за счет функции факторов транскрипции. Было показано, что Arabidopsis MPK6 действует, способствуя расщеплению и ядерной транслокации ORESARA3 (ORE3) / ETHYLENE INSENSITIVE2 (EIN2), белка, участвующего в старении листьев, опосредованном множеством факторов, включая ABA, этилен, MeJA, возраст и темноту. Освободившийся С-конец ORE3 / EIN2 (CEND) стабилизировал EIN3, фактор транскрипции, который ускоряет вызванное MeJA старение листьев [118].Интересно, что Arabidopsis MAPKKK MEKK1 был идентифицирован как ДНК-связывающий белок, который непосредственно регулирует фактор транскрипции WRKY53, способствующий старению [119].
4.3. Другие компоненты в передаче сигналов о старении
Помимо каскада киназ MAP, другие сигнальные системы, такие как передача сигналов, связанных с кальцием (Ca2 +), также участвуют в регуляции старения листьев. Было показано, что ингибитор-1 Arabidopsis Bax (AtBI1) замедляет вызванное MeJA старение листьев путем подавления [Ca 2+ ] цит-зависимой активации MPK6 [120].Кальциневрин B-подобная взаимодействующая протеинкиназа 14 (CIPK14) взаимодействовала и фосфорилировала оцДНК-связывающий белок WHIRLY1 у Arabidopsis [121]. WHIRLY1 был идентифицирован как белок, расположенный в пластидном ядре, который играет роль в регуляции старения листьев [122]. Фосфорилирование WHIRLY1 с помощью CIPK14 приводит к увеличению накопления белка в ядре и усилению связывания с промотором регулирующего старение фактора транскрипции WRKY53 [121].
Фосфатаза 2C-типа протеинфосфатаза, SENESCENCE-SUPPRESSED PROTEIN PHOSPHATASE (SSPP), была идентифицирована как сигнальный компонент, функционирующий ниже LRR-RLK AtSARK.Было обнаружено, что SSPP способен взаимодействовать с цитоплазматическим доменом AtSARK и дефосфорилировать его. Сверхэкспрессия SSPP может спасти преждевременное старение листьев и изменения гормональных ответов, вызванные AtSARK [123]. Другая протеинфосфатаза 2C, SAG113, была идентифицирована как прямой ген-мишень фактора транскрипции AtNAP. SAG113 был охарактеризован как негативный регулятор передачи сигналов ABA, который специфически участвует в контроле потери воды во время процесса старения листьев [24,27].
5. Выводы и перспективы
Передача сигнала является критической фазой программы старения листа, определяющей активацию нижестоящей регуляторной сети старения. За последние два десятилетия понимание регуляции старения листьев достигло многого, особенно в области гормональной регуляции и регуляции транскрипции старения листьев. Была проделана большая работа по восприятию и передаче сигналов, и было идентифицировано большое количество сигнальных компонентов ().Однако охарактеризовано лишь очень ограниченное количество сигнальных каскадов, таких как каскад AtSARK-SSPP [123], каскад MKK9-MPK6 [90] у Arabidopsis и каскад ZmMEK1-ZmSIMK1 у кукурузы [117]. Необходима большая работа для выяснения сложности и деталей регуляторной сети старения листьев между сигналами и факторами транскрипции путем идентификации большего количества сигнальных белков и, возможно, что более важно, путем установления связей между различными компонентами, которые уже были идентифицированы.Некоторые из критических вопросов передачи сигналов о старении листьев остаются нерешенными. Например, что является сигналом старения, как сигнал / сигнал старения воспринимается / воспринимается растительными клетками и как он преобразуется в факторы транскрипции для изменения экспрессии генов.
Сигнальные компоненты, идентифицированные в процессе старения листа.
Доступные идентифицированные компоненты передачи сигнала могут предоставить полезную платформу для разработки различных стратегий или систем для управления старением листьев для сельскохозяйственного производства.На основе хорошо изученного гомонального контроля и транскрипционного контроля старения листьев были разработаны технологии управления возрастом для замедления старения листьев, повышения устойчивости к стрессу и повышения урожайности сельскохозяйственных культур. Система SAG12-IPT использовалась для эффективной задержки старения листьев у более чем 20 видов растений, а технология на основе фактора транскрипции NAP также применялась более чем к десятку видов растений [45]. Было установлено, что ряд белков, регулирующих старение, в каскаде MAP-киназ также участвует в стрессовых ответах [14,90].Генетическая манипуляция сигнальными компонентами на стыках перекрестных переговоров между старением и реакциями на стресс может потенциально приводить к повышению устойчивости к стрессу и замедлению старения одновременно. С другой стороны, по мере выявления все большего количества сигнальных компонентов регуляторной сети старения листьев и раскрытия более полных сигнальных путей, ожидается, что для улучшения сельского хозяйства будут разработаны более специфичные и даже более эффективные стратегии манипулирования старением.
Благодарности
Авторы выражают благодарность сотрудникам лаборатории Гуо в Исследовательском институте табака Китайской академии сельскохозяйственных наук (CAAS) за обсуждения.
Вклад авторов
Концептуализация, Y.G .; письменная — подготовка оригинального черновика, S.A .; написание — просмотр и редактирование, Ю.Г .; наблюдение, Ю.Г .; администрация проекта, Ю.Г .; привлечение финансирования, Ю.Г.
Финансирование
Эта работа финансировалась Фондом фундаментальных исследований Китайской академии сельскохозяйственных наук (2013ZL024) и Программой инноваций в области сельскохозяйственных наук и технологий (ASTIP-TRIC02).
Конфликт интересов
Авторы заявляют об отсутствии конфликта интересов.
Ссылки
1. Солтабаева А., Шривастава С., Курманбаева А., Бектурова А., Флюхр Р., Саги М. Раннее старение старых листьев Atxdh2, выращенного с низким содержанием нитратов, выявляет роль катаболизма пуринов в поставке азота. Plant Physiol. 2018; 178: 1027–1044. DOI: 10.1104 / стр 18.00795. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 2. Буэт А., Коста М.Л., Мартинес Д.Э., Гьямет Х.Дж. Распад белка хлоропластов в стареющих листьях: протеазы и литические компартменты.Передний. Plant Sci. 2019; 10: 747. DOI: 10.3389 / fpls.2019.00747. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 3. Tamary E., Nevo R., Naveh L., Levin-Zaidman S., Kiss V., Savidor A., Levin Y., Eyal Y., Reich Z., Adam Z. Катаболизм хлорофилла предшествует изменениям в структуре и протеоме хлоропластов. во время старения листьев. Завод Директ. 2019; 3: e00127. DOI: 10.1002 / pld3.127. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 4. Майяр А., Дикелу С., Биллард В., Лайне П., Гарника М., Прудент М., Гарсиа-Мина Ж.-М., Ивин Ж.-К., Оурри А. Ремобилизация минеральных питательных веществ в листьях во время старения листьев и модуляция дефицитом питательных веществ. Передний. Plant Sci. 2015; 6: 317. DOI: 10.3389 / fpls.2015.00317. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 5. Хаве М., Мармань А., Шардон Ф., Маскло-Добресс К. Ремобилизация азота во время старения листьев: уроки от Arabidopsis до сельскохозяйственных культур. J. Exp. Бот. 2016; 68: 2513–2529. DOI: 10,1093 / jxb / erw365. [PubMed] [CrossRef] [Google Scholar] 6.Gortari F., Guiamet J.J., Cortizo S.C., Graciano C. Листовая ржавчина тополя снижает накопление сухой массы и внутреннюю рециркуляцию азота более заметно при низкой доступности азота в почве и снижает рост следующей весной. Tree Physiol. 2018; 39: 19–30. DOI: 10.1093 / treephys / tpy081. [PubMed] [CrossRef] [Google Scholar] 7. Куай Б., Чен Дж., Хёртенштайнер С. Биохимия и молекулярная биология распада хлорофилла. J. Exp. Бот. 2017; 69: 751–767. DOI: 10.1093 / jxb / erx322. [PubMed] [CrossRef] [Google Scholar] 8.Ву Х. Р., Ким Х. Дж., Лим П. О., Нам Х. Г. Старение листьев: системы и аспекты синамики. Анну. Rev. Plant Biol. 2019; 70: 347–376. DOI: 10.1146 / annurev-arplant-050718-095859. [PubMed] [CrossRef] [Google Scholar] 9. Али А., Гао Х., Го Ю. Инициирование, развитие и генетические манипуляции старением листьев. Методы Мол. Биол. 2018; 1744: 9–31. DOI: 10.1007 / 978-1-4939-7672-0_2. [PubMed] [CrossRef] [Google Scholar] 10. Хенсель Л.Л., Грбич В., Баумгартен Д.А., Бликкер А.Б. Процессы развития и возрастные процессы, влияющие на продолжительность жизни и старение фотосинтетических тканей у арабидопсиса .Растительная клетка. 1993; 5: 553–564. DOI: 10.1105 / tpc.5.5.553. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 11. Лим П.О., Ким Х.Дж., Нам Х.Г. Старение листьев. Анну. Rev. Plant Biol. 2007. 58: 115–136. DOI: 10.1146 / annurev.arplant.57.032905.105316. [PubMed] [CrossRef] [Google Scholar] 12. Джибран Р., Хантер Д., Дейквель П. Гормональная регуляция старения листьев посредством интеграции сигналов развития и стресса. Завод Мол. Биол. 2013; 82: 547–561. DOI: 10.1007 / s11103-013-0043-2. [PubMed] [CrossRef] [Google Scholar] 13.Schippers J.H.M., Jing H.C., Hille J., Dijkwel P.P. Онтогенетический и гормональный контроль старения листьев. Процесс старения. Растения. 2007. 26: 145–170. [Google Scholar] 14. Мацуока Д., Ясуфуку Т., Фуруя Т., Нанмори Т. Индуцибельная абсцизовой кислотой Arabidopsis MAPKKK, MAPKKK18 регулирует старение листьев посредством своей киназной активности. Завод Мол. Биол. 2015; 87: 565–575. DOI: 10.1007 / s11103-015-0295-0. [PubMed] [CrossRef] [Google Scholar] 15. Ким Дж., Чанг К., Такер М.Л. Чтобы состариться: регулирующая роль этилена и жасмоновой кислоты в старении.Передний. Plant Sci. 2015; 6:20. DOI: 10.3389 / fpls.2015.00020. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 16. Van der Graaff E., Schwacke R., Schneider A., Desimone M., Flugge U.-I., Kunze R. Транскрипционный анализ мембранных транспортеров Arabidopsis и гормональных путей во время развития и индуцированного старения листьев. Plant Physiol. 2006; 141: 776–792. DOI: 10.1104 / стр.106.079293. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 17. Цзин Х.С., Шипперс Дж. Х., Хилле Дж., Дейквель П.П. Вызванное этиленом старение листьев зависит от возрастных изменений и генов OLD в Arabidopsis . J. Exp. Бот. 2005; 56: 2915–2923. DOI: 10.1093 / jxb / eri287. [PubMed] [CrossRef] [Google Scholar] 18. Бьюкенен-Волластон В., Пейдж Т., Харрисон Э., Бриз Э., Лим П.О., Нам Х.Г., Лин Дж.-Ф., Ву С.-Х., Свидзински Дж., Ишизаки К. и др. Сравнительный анализ транскриптома выявляет значительные различия в экспрессии генов и сигнальных путях между онтогенетическим старением и старением, вызванным темнотой / голоданием, у Arabidopsis .Плант Дж. 2005; 42: 567–585. DOI: 10.1111 / j.1365-313X.2005.02399.x. [PubMed] [CrossRef] [Google Scholar] 19. Ким Х.Дж., Хонг С.Х., Ким Ю.В., Ли И.Х., Джун Дж.Х., Фи Б.К., Рупак Т., Чон Х., Ли Й., Хонг Б.С. и др. Генный регуляторный каскад ассоциированных со старением факторов транскрипции NAC, активируемых ETHYLENE-INSENSITIVE2-опосредованной передачей сигналов старения листьев в Arabidopsis . J. Exp. Бот. 2014; 65: 4023–4036. DOI: 10.1093 / jxb / eru112. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 20.Грбич В., Блеккер А. Этилен регулирует время старения листьев у Arabidopsis . Плант Дж. 1995; 8: 595–602. DOI: 10.1046 / j.1365-313X.1995.8040595.x. [CrossRef] [Google Scholar] 22. Мори И.К., Мурата Ю., Ян Ю., Мунемаса С., Ван Й.-Ф., Андреоли С., Тириак Х., Алонсо Дж. М., Харпер Дж. Ф., Эккер Дж. Р. и др. CDPKs CPK6 и CPK3 участвуют в регуляции АБК анионо- и Ca (2 +) — проницаемых каналов замыкающих клеток S-типа и закрытия устьиц. PLoS Biol. 2006; 4: e327. DOI: 10.1371 / journal.pbio.0040327. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 23. Munemasa S., Muroyama D., Nagahashi H., Nakamura Y., Mori I.C., Murata Y. Регулирование передачи сигналов абсцизовой кислоты, опосредованной реактивными формами кислорода, в замыкающих клетках и засухоустойчивость с помощью глутатиона. Передний. Plant Sci. 2013; 4: 472. DOI: 10.3389 / fpls.2013.00472. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 24. Zhang K., Xia X., Zhang Y., Gan S.S. ABA-регулируемая и локализованная по Гольджи протеинфосфатаза контролирует потерю воды во время старения листьев у Arabidopsis .Плант Дж. 2012; 69: 667–678. DOI: 10.1111 / j.1365-313X.2011.04821.x. [PubMed] [CrossRef] [Google Scholar] 25. Чжао Ю., Чан З., Гао Дж., Син Л., Цао М., Ю К., Ху Ю., Ю Дж., Ши Х., Чжу Ю. и др. Рецептор ABA PYL9 способствует засухоустойчивости и старению листьев. Proc. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ. 2016; 113: 1949–1954. DOI: 10.1073 / pnas.1522840113. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 26. Чжао Ю., Гао Дж., Им Ким Дж., Чен К., Брессан Р.А., Чжу Ж.-К. Контроль использования воды растениями с помощью индукции старения и покоя АБК: упущенный урок эволюции.Physiol растительной клетки. 2017; 58: 1319–1327. DOI: 10.1093 / pcp / pcx086. [PubMed] [CrossRef] [Google Scholar] 27. Чжан К., Ган С.-С. Регуляторная цепь абсцизовой кислоты-фактора транскрипции AtNAP-SAG113-протеинфосфатазы 2С для контроля обезвоживания в стареющих листьях Arabidopsis . Plant Physiol. 2012; 158: 961–969. DOI: 10.1104 / стр.111.1. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 28. Планас-Риверола А., Гупта А., Бетегон-Пуце И., Бош Н., Ибаньес М., Каньо-Дельгадо А.И. Передача сигналов брассиностероидами в развитии растений и адаптации к стрессу.Разработка. 2019; 146: dev151894. DOI: 10.1242 / dev.151894. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 29. Хасан С.А., Хаят С., Ахмад А. Брассиностероиды защищают фотосинтетические механизмы от окислительного стресса, вызванного кадмием, у двух сортов томатов. Chemosphere. 2011; 84: 1446–1451. DOI: 10.1016 / j.chemosphere.2011.04.047. [PubMed] [CrossRef] [Google Scholar] 30. Саглам-Каг С. Влияние эпибрассинолида на старение листьев пшеницы. Biotechnol. Biotechnol. Оборудуйте. 2007; 21: 63–65.DOI: 10.1080 / 13102818.2007.10817415. [CrossRef] [Google Scholar] 31. Ли З., Пан X., Гуо X., Фан К., Линь В. Физиологический и транскриптомный анализ раннего старения листьев для мутантного риса ospls1 ( Oryza sativa L.) на стадии заполнения зерна. Int. J. Mol. Sci. 2019; 20: 1098. DOI: 10.3390 / ijms20051098. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 32. Федина Е., Ярин А., Мухитова Ф., Блуфард А., Чечеткин И. Изменения липидного состава листьев Pisum sativum L. под действием брассиностероидов.во время старения. Стероиды. 2017; 117: 25–28. DOI: 10.1016 / j.steroids.2016.10.009. [PubMed] [CrossRef] [Google Scholar] 33. Wasternack C., Hause B. Jasmonates: биосинтез, восприятие, передача сигналов и действие в ответ на стресс, рост и развитие растений. Обновление обзора 2007 года в анналах ботаники. Анна. Бот. 2013; 111: 1021–1058. DOI: 10.1093 / aob / mct067. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 34. Уэда Дж., Като Дж. Стимулирующий эффект метилжасмоната на старение листьев овса на свету.Z. Pflanzenphysiol. 1981; 103: 357–359. DOI: 10.1016 / S0044-328X (81) 80134-1. [CrossRef] [Google Scholar] 35. He Y., Tang W., Swain J.D., Green A.L., Jack T.P., Gan S. Создание сети путей регуляции старения с использованием линий ловушек-энхансеров Arabidopsis . Plant Physiol. 2001; 126: 707–716. DOI: 10.1104 / стр. 126.2.707. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 36. He Y., Fukushige H., Hildebrand D.F., Gan S. Доказательства, подтверждающие роль жасмоновой кислоты в старении листьев Arabidopsis .Plant Physiol. 2002; 128: 876–884. DOI: 10.1104 / стр.010843. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 37. Бриз Э., Харрисон Э., Макхэтти С., Хьюз Л., Хикман Р., Хилл С., Киддл С., Ким Ю.С., Пенфолд С.А., Дженкинс Д. и др. Временное профилирование с высоким разрешением транскриптов во время старения листа Arabidopsis показывает четкую хронологию процессов и регуляции. Растительная клетка. 2011; 23: 873–894. DOI: 10.1105 / tpc.111.083345. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 38.Jung C., Lyou S.H., Yeu S., Kim M.A., Rhee S., Kim M., Lee J.S., Choi Y.D., Cheong J.-J. Скрининг на основе микрочипов чувствительных к жасмонату генов в Arabidopsis thaliana . Rep клетки растений 2007; 26: 1053-1063. DOI: 10.1007 / s00299-007-0311-1. [PubMed] [CrossRef] [Google Scholar] 39. Сяо С., Дай Л., Лю Ф., Ван З., Пэн В., Се Д. COS1: Нечувствительный к коронатину Arabidopsis супрессор , необходимый для регуляции опосредованной жасмонатом защиты и старения растений. Растительная клетка.2004. 16: 1132–1142. DOI: 10.1105 / tpc.020370. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 40. Тан X.-L., Fan Z.-Q., Shan W., Yin X.-R., Kuang J.-F., Lu W.-J., Chen J.-Y. Ассоциация BrERF72 с индуцированным метилжасмонатом старением листьев китайской цветущей капусты посредством активации генов, связанных с биосинтезом JA. Hortic. Res. 2018; 5: 22. DOI: 10.1038 / s41438-018-0028-z. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 41. Флот А.К., Демпси Д.М.А., Клессиг Д.Ф. Салициловая кислота, многогранный гормон для борьбы с болезнями.Анну. Rev. Phytopathol. 2009; 47: 177–206. DOI: 10.1146 / annurev.phyto.050908.135202. [PubMed] [CrossRef] [Google Scholar] 42. Besseau S., Li J., Palva E.T. WRKY54 и WRKY70 сотрудничают в качестве негативных регуляторов старения листьев у Arabidopsis thaliana . J. Exp. Бот. 2012; 63: 2667–2679. DOI: 10.1093 / jxb / err450. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 43. Го П., Ли З., Хуан П., Ли Б., Фанг С., Чу Дж., Го Х. Трехсторонняя петля амплификации, включающая фактор транскрипции WRKY75, салициловую кислоту и активные формы кислорода, ускоряет старение листьев.Растительная клетка. 2017; 29: 2854–2870. DOI: 10.1105 / tpc.17.00438. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 44. Ган С., Амасино Р.М. Подавление старения листьев за счет ауторегулируемого производства цитокинина. Наука. 1995; 270: 1986–1988. DOI: 10.1126 / science.270.5244.1986. [PubMed] [CrossRef] [Google Scholar] 45. Го Ю., Ган С.С. Трансляционные исследования старения листьев для повышения продуктивности и качества растений. J. Exp. Бот. 2014; 65: 3901–3913. DOI: 10.1093 / jxb / eru248. [PubMed] [CrossRef] [Google Scholar] 46.Мереитц Э. Б., Джанфагна Т., Хуанг Б. Накопление белка в листьях и корнях, связанное с улучшенной устойчивостью к засухе ползучей полевицы, экспрессирующей ген ipt для синтеза цитокининов. J. Exp. Бот. 2011; 62: 5311–5333. DOI: 10.1093 / jxb / err166. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 47. Rivero R.M., Gimeno J., Van Deynze A., Walia H., Blumwald E. Повышенный синтез цитокининов в растениях табака, экспрессирующих PSARK :: IPT, предотвращает деградацию фотосинтетических белковых комплексов во время засухи.Physiol растительной клетки. 2010; 51: 1929–1941. DOI: 10.1093 / pcp / pcq143. [PubMed] [CrossRef] [Google Scholar] 49. Kim H.J., Ryu H., Hong S.H., Woo H.R., Lim P.O., Lee I.C., Sheen J., Nam H.G., Hwang I. Цитокинин-опосредованный контроль долголетия листьев с помощью AHK3 посредством фосфорилирования ARR2 в Arabidopsis . Proc. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ. 2006; 103: 814–819. DOI: 10.1073 / pnas.0505150103. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 50. Бреннер В.Г., Романов Г.А., Коллмер И., Беркл Л., Шмуллинг Т.Гены немедленного-раннего и замедленного цитокининового ответа Arabidopsis thaliana , идентифицированные с помощью полногеномного профилирования экспрессии, выявляют новые чувствительные к цитокинину процессы и предполагают действие цитокининов через транскрипционные каскады. Плант Дж. 2005; 44: 314–333. DOI: 10.1111 / j.1365-313X.2005.02530.x. [PubMed] [CrossRef] [Google Scholar] 51. Али А., Гао Х., Го Ю. Старение растений. Humana Press; Нью-Йорк, Нью-Йорк, США: 2018. Инициирование, развитие и генетические манипуляции старением листьев; стр.9–31. [Google Scholar] 52. Но Ю.С., Амасино Р.М. Идентификация промоторной области, ответственной за специфическую для старения экспрессию SAG12. Завод Мол. Биол. 1999; 41: 181–194. DOI: 10,1023 / А: 1006342412688. [PubMed] [CrossRef] [Google Scholar] 53. Ким Дж.И., Мерфи А.С., Бэк Д., Ли С.В., Юн Д.Дж., Брессан Р.А., Нарасимхан М.Л. Сверхэкспрессия YUCCA6 демонстрирует функцию ауксина в задержке старения листьев у Arabidopsis thaliana . J. Exp. Бот. 2011; 62: 3981–3992. DOI: 10.1093 / jxb / err094.[Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 54. Кант С., Би Ю.-М., Чжу Т., Ротштейн С.Дж. SAUR39, небольшой ген РНК, повышающей ауксин, действует как негативный регулятор синтеза и транспорта ауксина в рисе. Plant Physiol. 2009; 151: 691–701. DOI: 10.1104 / стр.109.143875. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 55. Эллис К.М., Нагпал П., Янг Дж. К., Хаген Г., Гилфойл Т. Дж., Рид Дж. У. AUXIN RESPONSE FACTOR1 и AUXIN RESPONSE FACTOR2 регулируют старение и опадение цветочных органов у Arabidopsis thaliana .Разработка. 2005; 132: 4563. DOI: 10.1242 / dev.02012. [PubMed] [CrossRef] [Google Scholar] 56. Lim P.O., Lee I.C., Kim J., Kim H.J., Ryu J.S., Woo H.R., Nam H.G. Фактор ответа на ауксин 2 (ARF2) играет важную роль в регулировании опосредованного ауксином долголетия листьев. J. Exp. Бот. 2010; 61: 1419–1430. DOI: 10.1093 / jxb / erq010. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 57. Куирино Б.Ф., Норманли Дж., Амасино Р.М. Разнообразный диапазон активности генов во время старения листьев Arabidopsis thaliana включает патоген-независимую индукцию генов, связанных с защитой.Завод Мол. Биол. 1999. 40: 267–278. DOI: 10,1023 / А: 1006199932265. [PubMed] [CrossRef] [Google Scholar] 58. Ахарони Н., Андерсон Дж. Д., Либерман М. Производство и действие этилена на стареющие диски листьев: влияние индолуксусной кислоты, кинетина, иона серебра и диоксида углерода. Plant Physiol. 1979; 64: 805–809. DOI: 10.1104 / стр.64.5.805. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 59. Родригес К., Ванденберге Л.П.Д., де Оливейра Дж., Соккол К.Р. Новые перспективы производства гибберелловой кислоты: обзор.Крит. Rev. Biotechnol. 2012; 32: 263–273. DOI: 10.3109 / 07388551.2011.615297. [PubMed] [CrossRef] [Google Scholar] 60. Сан Т.-П., Габлер Ф. Молекулярный механизм передачи сигналов гиббереллина в растениях. Анну. Rev. Plant Biol. 2004. 55: 197–223. DOI: 10.1146 / annurev.arplant.55.031903.141753. [PubMed] [CrossRef] [Google Scholar] 61. Флетчер Р.А., Осборн Д.Дж. Регулирование синтеза белков и нуклеиновых кислот гиббереллином во время старения листьев. Природа. 1965; 207: 1176–1177. DOI: 10.1038 / 2071176a0. [CrossRef] [Google Scholar] 62.Ли Дж. Р., Ю. К., Вэй Дж. Р., Ма К., Ван Б. К., Ю. Д. Гиббереллин замедляет деградацию хлорофилла во время старения Paris polyphylla. Биол. Растение. 2010; 54: 395–399. DOI: 10.1007 / s10535-010-0072-5. [CrossRef] [Google Scholar] 63. Ю. К., Ван Ю., Вэй Дж., Ма К., Ю. Д., Ли Дж. Повышение урожайности и качества корневища Paris polyphylla за счет замедления старения надземных частей, вызванного гибберелловой кислотой. Завод сигнал. Behav. 2009; 4: 413–415. DOI: 10.4161 / psb.4.5.8268. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 64.Го Ю. К системному биологическому пониманию старения листьев. Завод Мол. Биол. 2013; 82: 519–528. DOI: 10.1007 / s11103-012-9974-2. [PubMed] [CrossRef] [Google Scholar] 65. Сехон Р.С., Саски К., Кумар Р., Флинн Б.С., Ло Ф., Бейсинджер Т.М., Акерман А.Дж., Брейцман М.В., Бриджес В.С., де Леон Н. и др. Интегрированный анализ в масштабе генома выявляет новые гены и сети, лежащие в основе старения кукурузы. Растительная клетка. 2019; 31: 1968–1989. DOI: 10.1105 / TPC.18.00930. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 66.Го Ю., Ган С. Старение листьев: сигналы, исполнение и регулирование. Curr. Верхний. Dev. Биол. 2005. 71: 83–112. DOI: 10.1016 / S0070-2153 (05) 71003-6. [PubMed] [CrossRef] [Google Scholar] 67. Ли В., Го Ю.Ф. Транскриптом, факторы транскрипции и регуляция транскрипции старения листа. J. Bioinforma. Комп. Геномика. 2014; 1: 1–3. [Google Scholar] 68. Го Ю., Цай З., Ган С. Транскриптом старения листьев Arabidopsis . Plant Cell Environ. 2004. 27: 521–549. DOI: 10.1111 / j.1365-3040.2003.01158.Икс. [CrossRef] [Google Scholar] 69. Ву Х. Программирование старения листьев растений с временной и межорганической координацией транскриптома в Arabidopsis . Plant Physiol. 2016; 171: 452–467. DOI: 10.1104 / стр.15.01929. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 70. Балазаде С., Риано-Пачон Д.М., Мюллер-Робер Б. Факторы транскрипции, регулирующие старение листьев у Arabidopsis thaliana .Plant Biol. 2008; 10: 63–75. DOI: 10.1111 / j.1438-8677.2008.00088.x. [PubMed] [CrossRef] [Google Scholar] 71. Ли В., Ли X., Чао Дж., Чжан З., Ван В., Го Ю. Факторы транскрипции семейства NAC в табаке и их потенциальная роль в регулировании старения листьев. Передний. Plant Sci. 2018; 9: 1900. DOI: 10.3389 / fpls.2018.01900. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 72. Shah S.T., Pang C.Y., Hussain A., Fan S.L., Song M.Z., Zamir R., Yu S.X. Молекулярное клонирование и функциональный анализ генов семейства NAC, связанных со старением листьев и стрессами у Gossypium hirsutum L.Культ растительных клеток, тканей, органов. 2014; 117: 167–186. DOI: 10.1007 / s11240-014-0430-7. [CrossRef] [Google Scholar] 73. Ким Х.Дж., Нам Х.Г., Лим П.О. Регуляторная сеть факторов транскрипции NAC при старении листьев. Curr. Opin. Plant Biol. 2016; 33: 48–56. DOI: 10.1016 / j.pbi.2016.06.002. [PubMed] [CrossRef] [Google Scholar] 74. Guo Y., Gan S. AtNAP, фактор транскрипции семейства NAC, играет важную роль в старении листьев. Плант Дж. 2006; 46: 601–612. DOI: 10.1111 / j.1365-313X.2006.02723.x. [PubMed] [CrossRef] [Google Scholar] 75.Balazadeh S., Kwasniewski M., Caldana C., Mehrnia M., Zanor MI, Xue GP, Mueller-Roeber B. ORS1, H (2) O (2) -чувствительный фактор транскрипции NAC, контролирует старение в Arabidopsis thaliana . Мол. Растение. 2011; 4: 346–360. DOI: 10,1093 / mp / ssq080. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 76. Гарапати П., Сюэ Г.-П., Мунне-Бош С., Балазаде С. Фактор транскрипции ATAF1 в Arabidopsis способствует старению путем прямой регуляции ключевого поддержания хлоропластов и транскрипционных каскадов старения.Plant Physiol. 2015; 168: 1122–1139. DOI: 10.1104 / стр.15.00567. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 77. Ву А., Аллу А.Д., Гарапати П., Сиддики Х., Дортай Х., Занор М.И., Асенси-Фабадо М.А., Мунне-Бош С., Антонио К., Тохге Т. и др. JUNGBRUNNEN1, фактор транскрипции NAC, реагирующий на активные формы кислорода, регулирует продолжительность жизни у арабидопсиса . Растительная клетка. 2012; 24: 482–506. DOI: 10.1105 / tpc.111.0 Краткий ответ: По мере того как лето переходит в осень, дни становятся короче и солнечного света становится все меньше.Это сигнал для листа, чтобы подготовиться к зиме и прекратить вырабатывать хлорофилл. Как только это происходит, зеленый цвет начинает тускнеть, и становятся видны красные, оранжевые и желтые оттенки. Красочные осенние листья. Фото Джереми Баукома (Flickr Creative Commons) Осенью деревья устраивают довольно впечатляющий показ мод. Листья, которые все лето были зелеными, начинают становиться ярко-красными, оранжевыми и желтыми. Но откуда эти цвета? Все начинается внутри листа.Листья имеют цвет из-за химических веществ, называемых пигментами, и в каждом листе есть четыре основных типа пигментов: Хлорофилл (зелень) Ксантофиллы (желтые) Каротиноиды (апельсины) Антоцианы (красные) Листья зеленые весной и летом, потому что именно тогда они производят много хлорофилла. Хлорофилл важен, потому что он помогает растениям получать энергию из солнечного света — процесс, называемый фотосинтезом. Летний солнечный свет заставляет листья производить больше хлорофилла. Но деревья очень чувствительны к изменениям в окружающей их среде. По мере того, как лето сменяется осенью, дни становятся короче и солнечного света становится все меньше. Это сигнал для листа, чтобы подготовиться к зиме и прекратить вырабатывать хлорофилл. Как только это происходит, зеленый цвет начинает тускнеть, и становятся видны красные, оранжевые и желтые оттенки. Более короткие осенние дни — это сигнал для деревьев, чтобы начать подготовку к зиме.Фото Тома Скарбека-Вазынски (Flickr Creative Commons) Погода также играет важную роль в изменении цвета. Осенью температуры становятся холоднее, а также больше
дождь и снег. Изменения этих погодных условий могут повлиять на то, насколько рано меняются листья и как долго они сохраняют свой красивый цвет. Экстремальные погодные изменения также могут повлиять на листья. Например, из-за засухи или ранних заморозков листья с деревьев опадут раньше, чем они успеют сменить цвет. Так что, если вы хотите знать, сколько цветов продержится этой осенью, обязательно следите за погодой! Стволовые клетки не только инициируют органы, но также могут способствовать формированию паттерна органов, по крайней мере, на верхушках побегов цветущих растений : классические микрохирургические эксперименты предполагают, что апикальная меристема побега способствует развитию адаксиальной стороны листа, т.е.е. верхняя сторона (Sussex, 1951, Nature 167: 651-652). «Sussex signal» был предложен как гипотетическая молекула, которая проходит от стволовых клеток верхушки побега к адаксиальному домену зарождающегося зачатка листа, чтобы способствовать образованию адаксиального домена (серая стрелка на рисунке 1). Однако химическая основа этого сигнала остается загадкой, как и его способ передачи и то, как он действует в зависимости от полярности листьев. Более ранние исследования официального Шанхайского института физиологии растений Китайской академии наук (CAS) показали, что ауксин участвует в полярности листьев (Ni et al., 1999, Cell Res. 9: 27-35). Следуя этому направлению, исследователи из группы JIAOYuling из Института генетики и биологии развития, CAS, вместе с коллегами из Калифорнийского технологического института и ENS-Lyon, показали существование временной зоны с низким содержанием ауксина на адаксиальной стороне, которая способствует адаксиальному развитию. Далее они обнаружили, что эта адаксиальная зона с низким содержанием ауксина является результатом транспорта ауксина от листьев к верхушке побега (красная стрелка на рисунке 1). Таким образом, это не положительный сигнал от стволовых клеток, а отход сигнальной молекулы от зачатков к стволовым клеткам, который передает информацию о полярности — противоположную тому, что обычно предполагается.Сигнал ауксина опосредуется чувствительным к ауксину фактором транскрипции MONOPTEROS (MP), конститутивная активация которого в адаксиальном домене способствует судьбе абаксиальных клеток. Это исследование дает объяснение гипотетическому сигналу Sussex о полярности листьев, полученной из меристемы. Эта работа была опубликована в Интернете в PNAS в Интернете Раннее издание 16 декабря 2014 г. ( DOI: 10.1073 / pnas.1421878112), с аспирантом Ци Цзиянем и докторантом Ван Ин как соавтором. -первые авторы.Это исследование было поддержано Национальной программой фундаментальных исследований Китая, Национальным фондом естественных наук Китая, Государственной ключевой лабораторией геномики растений и CAS. Рисунок 1. Концептуальная модель влияния на полярность транспорта ауксина от пазухи листа к SAM. (Изображение IGDB) 1. Ballaré CL, Sanchez RA, Scopel AL, Ghersa CM (1988) Морфологические реакции Datura-ferox L.саженцы на наличие соседей. Их взаимосвязь с микроклиматом полога . Oecologia 76: 288–293. 2. Pierik R, Visser EJW, De Kroon H, Voesenek LACJ (2003) Этилен требуется в to- bacco, чтобы успешно конкурировать с ближайшими соседями. Среда растительной клетки 26: 1229–1234. 3. Schmitt J, McCormac AC, Smith H (1995) Тест гипотезы адаптивной пластичности с использованием трансгенных и мутантных растений, лишенных фитохром-опосредованного удлинения ответов соседям.Am Nat 146: 937–953. 4. Франклин К.А. (2008) Избегание тени. Новый Фитол. 179: 930–944. 5. Vandenbussche F, Pierik R, Millenaar FF, Voesenek LACJ, Van Der Straeten D (2005) Выход из тени. Curr Opin Plant Biol 8: 462–468. 6. Martínez-García JF, et al. (2010) «Достижения в ботанических исследованиях», под редакцией Kader JC, Delseny M (Academic, New York), стр. 65–116. 7. Morgan DC, Smith H (1978) Взаимосвязь между фитохромом-фоторавновесием и развитием у выращенных на свету Chenopodium album L.Планта 142: 187–193. 8. Морган Д.К., О’Брайен Т., Смит Х. (1980) Быстрая фотомодуляция удлинения стебля у выращенных на свету Sinapis alba L. Planta 150: 95–101. 9. Балларе К.Л., Скопель А.Л., Санчес Р.А. (1990) Дальнее красное излучение, отраженное от соседних листьев: ранний сигнал о конкуренции в пологах растений. Наука 247: 329–332. 10. Балларе К.Л., Скопель А.Л., Санчес Р.А. (1991) Фотоуправление удлинением стебля у растений окрестностей: влияние скорости фотонного излучения в естественных условиях радиации. Среда растительных клеток 14: 57–65. 11. Pierik R, Whitelam GC, Voesenek LACJ, de Kroon H, Visser EJW (2004) Исследования Canopy на нечувствительном к этилену табаке определяют этилен как новый элемент в синем свете и сигнализирующих о растениях. Завод J 38: 310–319. 12. Kasperbauer MJ (1971) Спектральное распределение света в навесе табака и влияние качества дневного света на рост и развитие . Физиология растений 47: 775–778. 13.Ballaré CL, Sánchez RA, Scopel AL, Casal JJ, Ghersa CM (1987) Раннее обнаружение соседних растений по восприятию фитохромом спектральных изменений в отраженном солнечном свете. Среда растительных клеток 10: 551–557. 14. Гилберт И.Р., Джарвис П.Г., Смит Х. (2001) Сигнал близости и различия в избегании тени между ранними и поздними сукцессионными деревьями. Природа 411: 792–795. 15. Карабелли М., Сесса Г., Байма С., Морелли Г., Руберти I (1993) Гены Arabidopsis Athb-2 и -4 сильно индуцируются ярко-красным светом.Завод J 4: 469–479. 16. Sasidharan R, et al. (2010) Опосредованное качеством света удлинение черешка у арабского идопсиса во время избегания тени включает модификацию клеточной стенки с помощью ксилоглюкан-эндотрансглюкозилазы / гидролаз. Физиология растений 154: 978–990. 17. Степанова А.Н., и др. (2008) TAA1-опосредованный биосинтез ауксина важен для перекрестных помех гормонами и развития растений. Ячейка 133: 177–191. 18. Tao Y, et al. (2008) Быстрый синтез ауксина по новому триптофан-зависимому пути — — необходим для избегания тени у растений.Ячейка 133: 164–176. 19. Браам Дж. (2005) На связи: реакция растений на механические раздражители. Новый Фитол 165: 373–389. 20. Chehab EW, Yao C, Henderson Z, Kim S, Braam J (2012) Морфогенез , вызванный прикосновением Arabidopsis, опосредован жасмонатом и защищает от вредителей. Curr Biol 22: 701–706. 21. Cox MCH, Millenaar FF, Van Berkel YE, Peeters AJM, Voesenek LACJ (2003) Механизм завода . Вызванное погружением удлинение черешка у Rumex palustris зависит от гипонастического роста .Физиология растений 132: 282–291. 22. Evers JB, et al. (2007) Моделирование влияния локализованного соотношения красного: дальнего красного на кущение яровой пшеницы (Triticum aestivum) с использованием трехмерной виртуальной модели растения. New Phytol 176: 325–336. 23. Смит Х. (2000) Фитохромы и восприятие светового сигнала растениями — новый синтез . Nature 407: 585–591. 24. Pierik R, De Wit M, Voesenek LACJ (2011) Ускользание от стресса, обусловленное ростом: Conver- генов путей передачи сигнала, активируемых при воздействии двух различных стрессов окружающей среды .Новый Фитол. 189: 122–134. 25. Coutand C (2010) Механочувствительность и тигмоморфогенез, физиологическая и биомеханическая точки зрения. Plant Sci 179: 168–182. 26. Anten NPR, Casado-Garcia R, Nagashima H (2005) Влияние механического стресса и плотности растений на механические характеристики, рост и воспроизводство в течение всей жизни растений табака. Am Nat 166: 650–660. 27. Takahashi H, Suge H, Kato T (1991) Стимуляция роста с помощью вибрации с частотой 50 Гц в рисе и саженцах огурцов.Физиология растительных клеток 32: 729-732. 28. Джонсон К.А., Систранк М.Л., Полисенский Д.Х., Браам Дж. (1998) Arabidopsis thaliana re- спонов для механической стимуляции не требуют ETR1 или EIN2. Физиология растений 116: 643–649. 29. Polko JK, et al. (2012) Дифференциальный рост черешка у Arabidopsis thaliana, индуцированный этиленом, включает локальную переориентацию микротрубочек и рост клеток. Новый Фитол 193: 339–348. 30. Lorrain S, Allen T, Duek PD, Whitelam GC, Fankhauser C (2008) Ингибирование избегания тени, вызванное фитохромом, , включает деградацию факторов транскрипции bHLH , способствующих росту.Завод J 53: 312–323. 31. Leivar P, et al. (2008) Фактор PIF7, взаимодействующий с фитохромом Arabidopsis, вместе с PIF3 и PIF4, регулирует реакцию на длительный красный свет, модулируя уровни phyB . Растительная клетка 20: 337–352. 32. Kim GT, Tsukaya H, Uchimiya H (1998) Ген ROTUNDIFOLIA3 Arabidopsis thaliana кодирует нового члена семейства цитохрома P-450, который необходим для регулируемого полярного удлинения клеток листа .Гены Дев 12: 2381–2391. 33. Friml J, Wiśniewska J, Benková E, Mendgen K, Palme K (2002) Боковое перемещение регулятора потока ауксина PIN3 опосредует тропизм у Arabidopsis. Природа 415: 806–809. 34. Bleecker AB, Estelle MA, Somerville C, Kende H (1988) Нечувствительность к этилену, кон- , вызванная доминантной мутацией в Arabidopsis thaliana. Science 241: 1086–1089. 35. Park JH, et al. (2002) Нокаут-мутация в синтазе алленоксида приводит к стерильности самцов и дефектной трансдукции раневого сигнала у Arabidopsis из-за блокирования биосинтеза жасмоновой кислоты .Завод J 31: 1–12. 36. Staswick PE, Yuen GY, Lehman CC (1998) Сигнальные мутанты Jasmonate Arabidopsis чувствительны к почвенному грибку Pythium irregulare.Plant J 15: 747–754. 37. Xie DX, Feys BF, James S, Nieto-Rostro M, Turner JG (1998) COI1: ген Arabidopsis , необходимый для регулируемой жасмонатом защиты и фертильности. Science 280: 1091–1094. 38. Cao H, Glazebrook J, Clarke JD, Volko S, Dong X (1997) Ген NPR1 арабидопсиса , который контролирует системную приобретенную устойчивость, кодирует новый белок, содержащий повторов анкирина.Cell 88: 57–63. 39. Clouse SD, Langford M, McMorris TC (1996) Нечувствительный к брассиностероидам мутант в Arabidopsis thaliana обнаруживает множественные дефекты роста и развития. Завод Physiol 111: 671–678. 40. Millenaar FF, et al. (2005) Дифференциальный рост черешков у Arabi- dopsis, индуцированный этиленом. Анализ естественной изменчивости, кинетики ответа и регуляции. Физиология растений 137: 998–1008. 41. Абрамов М.Д., Магальяйнс П.Дж., Рам С.Дж. (2004) Обработка изображений с помощью ImageJ.Биофотон Инт 11: 36–41. 42. Vos J, et al. (2010) Функционально-структурное моделирование растений: новый универсальный инструмент в науке о сельскохозяйственных культурах . J Exp Bot 61: 2101–2115. 43. Хеммерлинг Р., Книмейер О., Ланверт Д., Курт В., Бак-Сорлин Г. (2008) Язык XL на основе правила и среда моделирования GroIMP, проиллюстрированная с помощью моделирования конкуренции деревьев . Функциональная биология растений 35: 739–750. 44. Mündermann L, Erasmus Y, Lane B, Coen E, Prusinkiewicz P (2005) Количественное моделирование развития Arabidopsis.Физиология растений 139: 960–968. 45. Untergasser A, et al. (2007) Primer3Plus, усовершенствованный веб-интерфейс для Primer3. Nucleic Acids Res 35 (выпуск веб-сервера): W71-4. 46. Ливак К.Дж., Шмитген Т.Д. (2001) Анализ данных относительной экспрессии генов с использованием количественной ПЦР в реальном времени и метода 2 (-ΔΔC (T)) . Методы 25: 402–408. 47. Irizarry RA, et al. (2003) Исследование, нормализация и обобщение данных об уровне зондов с высокой плотностью матрицы олигонуклеотидов.Биостатистика 4: 249–264. 48. Смит Г.К. (2004) Линейные модели и эмпирические байесовские методы для оценки дифференциальной экспрессии в экспериментах с микрочипами. Stat Appl Genet Mol Biol 3: Статья3. 49. Benjamini Y, Hochberg Y (1995) Контроль ложного обнаружения: практический и мощный подход к множественному тестированию. J Roy Stat Soc B Met 57: 289–300. 50. Maere S, Heymans K, Kuiper M (2005) BiNGO: плагин Cytoscape для оценки более представлений категорий генной онтологии в биологических сетях.Биоинформатика 21: 3448–3449. 51. Основная группа разработчиков R (2008) R: Язык и среда для статистических вычислений Вычисления (Фонд R для статистических вычислений, Вена). 14710 | www.pnas.org/cgi/doi/10.1073/pnas.1205437109 de Wit et al. слов и картинок Информация FALLLLL: Подробнее покупка. Crown’d: Подробнее Прочтите бесплатно или купите. Runs on Blood: Нарисовано для Всемирного экономического форума 2019 года в Давосе, Швейцария, резидентом * Уорреном Крэгхедом III.. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 78. Ян С.Д., Со П.Дж., Юн Х.К., Пак К.М. Фактор транскрипции NAC
Arabidopsis VNI2 интегрирует сигналы абсцизовой кислоты в процесс старения листьев через гены COR / RD. Растительная клетка. 2011; 23: 2155–2168. DOI: 10.1105 / tpc.111.084913. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 79. Zhang W.Y., Xu Y.C., Li W.L., Yang L., Yue X., Zhang X.S., Zhao X.Y. Транскрипционный анализ естественного старения листьев кукурузы. PLoS ONE. 2014; 9: e115617. DOI: 10.1371 / journal.pone.0115617. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 80.Ким Х. Дж., Пак Дж. Х., Ким Дж., Ким Дж. Дж., Хон С., Ким Дж., Ким Дж. Х., Ву Х. Р., Хён С., Лим П. О. и др. Изменяющиеся во времени генетические сети обнаруживают тройку NAC, которая негативно регулирует старение листьев у Arabidopsis . Proc. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ. 2018; 115: E4930 – E4939. DOI: 10.1073 / pnas.1721523115. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 81. Eulgem T., Rushton P.J., Robatzek S., Somssich I.E. Надсемейство факторов транскрипции растений WRKY. Trends Plant Sci. 2000. 5: 199–206.DOI: 10.1016 / S1360-1385 (00) 01600-9. [PubMed] [CrossRef] [Google Scholar] 82. Робацек С., Сомсич И.Э. Мишени регуляции AtWRKY6 во время старения растений и защиты от патогенов. Genes Dev. 2002; 16: 1139–1149. DOI: 10.1101 / gad.222702. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 83. Робацек С., Сомсич И.Э. Новый член семейства транскрипционных факторов Arabidopsis WRKY, AtWRKY6, связан как с процессами, связанными со старением, так и с защитными процессами. Плант Дж. 2001; 28: 123–133.DOI: 10.1046 / j.1365-313X.2001.01131.x. [PubMed] [CrossRef] [Google Scholar] 84. Мяо Ю., Лаун Т., Циммерманн П., Зентграф У. Мишени фактора транскрипции WRKY53 и его роль во время старения листьев у Arabidopsis . Завод Мол. Биол. 2004. 55: 853–867. DOI: 10.1007 / s11103-005-2142-1. [PubMed] [CrossRef] [Google Scholar] 85. Ву Х. Р., Ким Х. Дж., Нам Х. Г., Лим П. О. Старение и гибель листьев растений — регулирование с помощью нескольких уровней контроля и влияние на старение в целом. J. Cell Sci.2013; 126: 4823. DOI: 10.1242 / jcs.109116. [PubMed] [CrossRef] [Google Scholar] 86. Zhou X., Jiang Y., Yu D. Фактор транскрипции WRKY22 опосредует индуцированное темнотой старение листьев в Arabidopsis . Мол. Ячейки. 2011; 31: 303–313. DOI: 10.1007 / s10059-011-0047-1. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 87. Шараби-Швагер М., Самач А., Порат Р. Сверхэкспрессия активатора транскрипции CBF2 в Arabidopsis противодействует гормональной активации старения листьев. Завод сигнал.Behav. 2010; 5: 296–299. DOI: 10.4161 / psb.5.3.10739. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 88. Ву Х.Р., Ким Дж. Х., Ким Дж., Ким Дж., Ли Ю., Сон И.-Дж., Ким Дж .-Х., Ли Х.-Й., Нам Х. Г., Лим П.О. Фактор транскрипции RAV1 положительно регулирует старение листьев у Arabidopsis . J. Exp. Бот. 2010; 61: 3947–3957. DOI: 10.1093 / jxb / erq206. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 89. Хопкинс М., Тейлор К., Лю З., Ма Ф., Макнамара Л., Ван Т.-В., Томпсон Дж. Э. Регулирование и выполнение молекулярной разборки и катаболизма во время старения.Новый Фитол. 2007; 175: 201–214. DOI: 10.1111 / j.1469-8137.2007.02118.x. [PubMed] [CrossRef] [Google Scholar] 90. Zhou C., Cai Z., Guo Y., Gan S. Каскад митоген-активируемых протеинкиназ Arabidopsis , MKK9-MPK6, играет роль в старении листьев. Plant Physiol. 2009. 150: 167–177. DOI: 10.1104 / стр.108.133439. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 92. Шиу С.Х., Бликер А.Б. Рецептороподобные киназы из Arabidopsis образуют семейство монофилетических генов, родственных рецепторным киназам животных.Proc. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ. 2001; 98: 10763–10768. DOI: 10.1073 / pnas.181141598. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 93. Шиу С.Х., Бликер А.Б. Семейство генов рецептороподобных киназ растений: разнообразие, функция и передача сигналов. Sci. СТКЕ. 2001; 2001: re22. DOI: 10.1126 / stke.2001.113.re22. [PubMed] [CrossRef] [Google Scholar] 94. Уэлхадж А., Камински М., Миттаг М., Хамбек К. Рецептороподобная протеинкиназа HvLysMR1 ячменя ( Hordeum vulgare L.) индуцируется во время старения листьев и стресса тяжелых металлов.J. Exp. Бот. 2007. 58: 1381–1396. DOI: 10.1093 / jxb / erl304. [PubMed] [CrossRef] [Google Scholar] 95. Shiu S.-H., Karlowski W.M., Pan R., Tzeng Y.-H., Mayer K.F.X., Li W.-H. Сравнительный анализ семейства рецепторных киназ у Arabidopsis и риса. Растительная клетка. 2004. 16: 1220–1234. DOI: 10.1105 / tpc.020834. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 96. Гофф К.Е., Рамонелл К.М. Роль и регуляция рецепторных киназ в защите растений. Джин Регул. Syst. Биол. 2007; 1: 167–175.DOI: 10.1177 / 117762500700100015. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 97. Кок Дж. М., Ванустхейз В., Гауд Т. Передача сигналов киназы рецепторов у растений и животных: отдельные молекулярные системы с механистическим сходством. Curr. Opin. Cell Biol. 2002. 14: 230–236. DOI: 10.1016 / S0955-0674 (02) 00305-8. [PubMed] [CrossRef] [Google Scholar] 98. Вэй З., Ван Дж., Ян С., Сонг Ю. Идентификация и анализ экспрессии семейства генов LRR-RLK в томате ( Solanum lycopersicum ) Heinz 1706.Геном. 2015; 58: 121–134. DOI: 10.1139 / gen-2015-0035. [PubMed] [CrossRef] [Google Scholar] 99. Zan Y., Ji Y., Zhang Y., Yang S., Song Y., Wang J. Полногеномная идентификация, характеристика и анализ экспрессии генов протеинкиназ populus, богатых лейцином, рецепторов. BMC Genomics. 2013; 14: 318. DOI: 10.1186 / 1471-2164-14-318. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 100. Li X., Salman A., Guo C., Yu J., Cao S., Gao X., Li W., Li H., Guo Y. Идентификация и характеристика генов семейства LRR-RLK у картофеля показывают их участие в пептидный сигнал о решениях клеточной судьбы и ответах на биотический / абиотический стресс.Ячейки. 2018; 7: 120. DOI: 10,3390 / ячейки70. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 101. Hajouj T., Michelis R., Gepstein S. Клонирование и характеристика рецепторного гена протеинкиназы, связанного со старением. Plant Physiol. 2000; 124: 1305–1314. DOI: 10.1104 / стр.124.3.1305. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 102. Делаторре С.А., Коэн Ю., Лю Л., Пелег З., Блюмвальд Э. Регулирование активности промотора SARK гормонами и сигналами окружающей среды.Plant Sci. 2012; 193–194: 39–47. DOI: 10.1016 / j.plantsci.2012.05.005. [PubMed] [CrossRef] [Google Scholar] 103. Ли X.P., Gan R., Li P.L., Ma Y.Y., Zhang L.W., Zhang R., Wang Y., Wang N.N. Идентификация и функциональная характеристика гена киназы, подобного рецептору, с богатым лейцином повтора, который участвует в регуляции старения листьев сои. Завод Мол. Биол. 2006; 61: 829–844. DOI: 10.1007 / s11103-006-0052-5. [PubMed] [CrossRef] [Google Scholar] 104. Сюй Ф., Мэн Т., Ли П., Ю Ю., Цуй Ю., Ван Ю., Гонг К., Ван Н. Киназа с двойной специфичностью, GmSARK, и ее гомолог Arabidopsis , AtSARK, регулируют старение листьев за счет синергетического действия ауксина и этилена. Plant Physiol. 2011; 157: 2131–2153. DOI: 10.1104 / стр.111.182899. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 105. Ли П., Ян Х., Лю Г., Ма В., Ли К., Хо Х., Хэ Дж., Лю Л. PpSARK регулирует старение мха и толерантность к соли с помощью пути, связанного с АБК. Int. J. Mol. Sci. 2018; 19: 2609. DOI: 10.3390 / ijms19092609.[Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 106. Lee I.C., Hong S.W., Whang S.S., Lim P.O., Nam H.G., Koo J.C. Возрастозависимое действие ABA-индуцируемой рецепторной киназы RPK1 как положительного регулятора старения в листьях Arabidopsis . Physiol растительной клетки. 2011; 52: 651–662. DOI: 10.1093 / pcp / pcr026. [PubMed] [CrossRef] [Google Scholar] 107. Li X., Ahmad S., Ali A., Guo C., Li H., Yu J., Zhang Y., Gao X., Guo Y. Характеристика рецептороподобной киназы 4 соматического эмбриогенеза как негативного регулятора листа старение Arabidopsis .Ячейки. 2019; 8:50. DOI: 10,3390 / ячейки8010050. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 108. Брандт Б., Корецепторные киназы Hothorn M. SERK. Curr. Биол. 2016; 26: R225 – R226. DOI: 10.1016 / j.cub.2015.12.014. [PubMed] [CrossRef] [Google Scholar] 109. Ху К., Чжу Ю., Цуй Ю., Ченг К., Лян В., Вэй З., Чжу М., Инь Х., Цзэн Л., Сяо Ю. и др. Группа рецепторных киназ важна для передачи сигналов CLAVATA для поддержания гомеостаза стволовых клеток. Nat. Растения. 2018; 4: 205–211. DOI: 10.1038 / s41477-018-0123-z.[PubMed] [CrossRef] [Google Scholar] 110. Цуй Ю., Ху К., Чжу Ю., Ченг К., Ли Х., Вэй З., Сюэ Л., Лин Ф., Ши Х., Йи Дж. И др. Киназы рецептора CIK определяют спецификацию клеточной судьбы во время раннего развития пыльника у Arabidopsis . Растительная клетка. 2018; 30: 2383. DOI: 10.1105 / tpc.17.00586. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 111. Цзэн Д.-Д., Ян Ч.-К., Цинь Р., Аламин М., Юэ Э.-К., Цзинь X.-Л., Ши Ч.-Х. Вставка гуанина в OsBBS1 приводит к раннему старению листьев и чувствительности к солевому стрессу у риса ( Oryza sativa L.) Plant Cell Rep.2018; 37: 933–946. DOI: 10.1007 / s00299-018-2280-у. [PubMed] [CrossRef] [Google Scholar] 112. Чжан М., Су Дж., Чжан Ю., Сюй Дж., Чжан С. Передача эндогенных и экзогенных сигналов: каскады MAPK в росте и защите растений. Curr. Opin. Plant Biol. 2018; 45: 1–10. DOI: 10.1016 / j.pbi.2018.04.012. [PubMed] [CrossRef] [Google Scholar] 113. Берберих Т., Сано Х., Кусано Т. Участие MAP-киназы, ZmMPK5, в старении и восстановлении кукурузы после низкотемпературного стресса. Мол. Genet Genet. MGG.1999; 262: 534–542. DOI: 10.1007 / s004380051115. [PubMed] [CrossRef] [Google Scholar] 114. Ван С.-Х., Лим Дж.-Х., Ким С.-С., Чо С.-Х., Ю С.-К., Ко Х.-Дж., Сакураба Ю., Пэк Н.- С. Мутация SPOTTED LEAF3 (SPL3) нарушает передачу сигналов, реагирующих на абсцизовую кислоту, и задерживает старение листьев у риса. J. Exp. Бот. 2015; 66: 7045–7059. DOI: 10.1093 / jxb / erv401. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 115. Тан Д., Иннес Р. В. Сверхэкспрессия киназодефицитной формы гена EDR1 усиливает устойчивость к мучнистой росе и индуцированное этиленом старение у арабидопсиса .Плант Дж. 2002; 32: 975–983. DOI: 10.1046 / j.1365-313X.2002.01482.x. [PubMed] [CrossRef] [Google Scholar] 116. Фрай К.А., Танг Д., Иннес Р.В. Отрицательная регуляция защитных ответов у растений консервативной киназой MAPKK. Proc. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ. 2001. 98: 373–378. DOI: 10.1073 / pnas.98.1.373. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 117. Ли Ю., Чанг Ю., Чжао К., Ян Х., Рен Д. Экспрессия неактивного ZmMEK1 вызывает накопление салициловой кислоты и зависимое от салициловой кислоты старение листьев.J. Integr. Plant Biol. 2016; 58: 724–736. DOI: 10.1111 / jipb.12465. [PubMed] [CrossRef] [Google Scholar] 118. Zhang Y., Liu J., Chai J., Xing D. Митоген-активированная протеинкиназа 6 опосредует ядерную транслокацию ORE3, чтобы способствовать экспрессии гена ORE9 при старении листьев, индуцированном метилжасмонатом. J. Exp. Бот. 2015; 67: 83–94. DOI: 10.1093 / jxb / erv438. [PubMed] [CrossRef] [Google Scholar] 119. Miao Y., Laun T.M., Smykowski A., Zentgraf U. Arabidopsis MEKK1 может быть коротким путем: он может напрямую взаимодействовать с связанным со старением транскрипционным фактором WRKY53 на уровне белка и может связываться с его промотором.Завод Мол. Биол. 2007; 65: 63–76. DOI: 10.1007 / s11103-007-9198-z. [PubMed] [CrossRef] [Google Scholar] 120. Yue H., Nie S., Xing D. Сверхэкспрессия Arabidopsis Bax ингибитор-1 задерживает вызванное метилжасмонатом старение листьев, подавляя активацию MAP-киназы 6. J. Exp. Бот. 2012; 63: 4463–4474. DOI: 10.1093 / jxb / ers122. [PubMed] [CrossRef] [Google Scholar] 121. Ren Y., Li Y., Jiang Y., Wu B., Miao Y. Фосфорилирование WHIRLY1 с помощью CIPK14 сдвигает его локализацию и двойные функции в Arabidopsis .Мол. Растение. 2017; 10: 749–763. DOI: 10.1016 / j.molp.2017.03.011. [PubMed] [CrossRef] [Google Scholar] 122. Miao Y., Jiang J., Ren Y., Zhao Z. Одноцепочечный ДНК-связывающий белок WHIRLY1 репрессирует экспрессию WRKY53 и задерживает старение листьев в зависимости от стадии развития у Arabidopsis . Plant Physiol. 2013. 163: 746–756. DOI: 10.1104 / стр.113.223412. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 123. Сяо Д., Цуй Ю., Сюй Ф., Сюй X., Гао Г., Ван Ю., Го З., Ван Д., Ван Н.N. ПОДАВЛЕННАЯ ЧУВСТВИТЕЛЬНОСТЬЮ БЕЛКОВАЯ ФОСФОТАЗА напрямую взаимодействует с цитоплазматическим доменом КИНАЗЫ, СВЯЗАННОЙ С СМЫСЛОМ, и отрицательно регулирует старение листьев в Arabidopsis . Plant Physiol. 2015; 169: 1275–1291. DOI: 10.1104 / стр.15.01112. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] Почему листья меняют цвет?
Ауксин как суссексский сигнал, определяющий полярность листьев — Институт генетики и биологии развития Китайской академии наук
(PDF) Обнаружение соседей растений через касание кончиков листьев предшествует сигналам фитохромов
ЛИСТ И СИГНАЛ
Работа, сделанная во время карантина 2020 художниками в Шарлоттсвилле, штат Вирджиния, США
Рисунки с Всемирного экономического форума 2019 в Давосе, Свитерленд
WEF — это ежегодное мероприятие, на котором лидеры бизнеса, грязные патриции и правительственные чиновники обсуждают будущее капитализма и способы наилучшего увеличения прибыли для себя и своих рыночных хозяев.
* Самопровозглашенные и работающие на дому.
40 страниц, цветное, 4,25 x 5,5 дюйма5 $ + доставка. Больше информации | Купить
ВЛАЖНАЯ КРАСКА, рассказ
, W. Craghead, 16 страниц, 4,25 x 5,5 дюйма, цветная Страницы расположены в случайном порядке, чтобы образовать десятки возможных историй.Каждая книга индивидуальна. Версия этой истории была опубликована в C’Est Bon Kultur. Книжный дебют на SPX 2019
$ 1 + доставка
Купить
Умный Нарвал: история происхождения
, Ginger Craghead-Way, 4,25 x 5,5 дюйма, 8 страниц, черно-белое
Узнайте происхождение союзника Veggie Team Умного Нарвала. Часть вселенной Veggie Team
$ 1 + доставка
Купить
Veggie Team issue 4
, Ginger Craghead-Way, 8-страничная книга, 8.5 x 5,5 дюйма, черно-белое
Еще приключения Кэрри и Барри, и они сражаются с врагами! Часть вселенной Veggie Team
$ 1 + доставка
Купить
Veggie Team issue 3
, Ginger Craghead-Way, 8-страничная книга, 8,5 x 5,5 дюйма, черно-белая
Еще приключения Кэрри и Барри. Часть вселенной Veggie Team
$ 1 + доставка
Купить
Veggie Team, выпуск 1 и 2
, Ginger Craghead-Way, 8-страничная книга, 8.5 x 5,5 дюйма, черно-белое
Познакомьтесь с Кэрри и Барри, которые веселятся и веселятся. Часть вселенной Veggie Team
$ 1 + доставка
Купить
КАК ВЕЗДЕ БЫТЬ
Комиксы по поэзии.
У. Крэгхеда. Книга на 90 страниц, цветная обложка, интерьер черно-белый
По поэзии Гийома Аполлинера
15 $ + доставка
Купить
TRUMPTRUMP: SOTU 2018
по W.Крэгхед. 52-страничная книга, 4,25 x 4,25 дюйма, цветная
Обращение Дональда Трампа к Конгрессу США о положении страны в прямом эфире от 30 января 2018 г.
Нарисовано во время просмотра, опубликовано в прямом эфире в инстаграмме, собрано в этой книге.
Также смотрите коллекции TRUMPTRUMP.biz и TRUMPTRUMP.$ 3 + доставка
Купить
Double Nickels Forever
Трофейная дань памяти Minutemen’s Double Nickels на альбоме Dime.На двойной LP 58 художников нарисовали 45 песен — 182 страницы нарисованного эконо.
9,95 $ + доставка
Купить
Дистанционные сигналы, регулирующие устьичную проводимость и рост листьев у растений томата (Lycopersicon esculentum), подвергнутых частичной сушке корневой зоны на JSTOR
АбстрактныйРастения томата (Lycopersicon esculentum Mill. Сорт Ailsa Craig) выращивали с корнями, разделенными между двумя столбиками почвы. После укоренения растений воду ежедневно подавали в одну (частичная сушка корневой зоны — PRD) или обе (хорошо поливная контрольная — WW) колонки.При обработке PRD из другой колонки откачивали воду, чтобы подвергнуть некоторые корни воздействию высыхающей почвы. Состояние воды в почве и растениях контролировалось ежедневно и в течение суток. За 8 дней обработки не было различий в потенциале воды в листьях (лист), даже несмотря на то, что содержание влаги в почве в верхних 6 см (θ) сухого столба при обработке PRD снизилось до 70%. Устьичная проводимость (gs) растений PRD снижалась (по сравнению с растениями WW), когда θ сухого столба уменьшалось на 45%. Такое закрытие совпало с повышенным pH ксилемного сока и не требовало увеличения концентрации абсцизовой кислоты (ABA) ксилемного сока ([X-ABA]).Выделение этилена в отделенных листочках растений PRD увеличивалось, когда θ в сухой колонке уменьшалось на 55%, одновременно с уменьшением удлинения листьев. Физиологическое значение усиленного выделения этилена в PRD-растениях было исследовано с использованием трансгенного томата (ACO1AS) с низким уровнем продуцирования этилена, индуцированного стрессом. В ответ на PRD, ACO1AS и растения дикого типа показали сходные pH ксилемного сока, [X-ABA] и gs, но растения ACO1AS не показали ни усиленного выделения этилена, ни значительного снижения удлинения листьев.Совместное использование генетических технологий для сокращения производства этилена и агрономических технологий для поддержания водного статуса (например, PRD) может поддерживать рост растений в условиях, при которых урожайность в противном случае была бы значительно снижена.
Информация о журналеЦелью JXB является публикация рукописей высочайшего качества, в которых рассматриваются вопросы, представляющие широкий интерес в биологии растений. Мы приветствуем рукописи, которые определяют фундаментальные механизмы, в том числе лежащие в основе улучшения растений для устойчивого производства продуктов питания, топлива и возобновляемых материалов.Мы делаем упор на молекулярную физиологию, молекулярную генетику и физиологию окружающей среды, и мы поощряем интегративные подходы с использованием передовых технологий, системной биологии и синтетической биологии.
Информация об издателеOxford University Press — это отделение Оксфордского университета. Издание во всем мире способствует достижению цели университета в области исследований, стипендий и образования. OUP — крупнейшая в мире университетская пресса с самым широким присутствием в мире.В настоящее время он издает более 6000 новых публикаций в год, имеет офисы примерно в пятидесяти странах и насчитывает более 5500 сотрудников по всему миру. Он стал известен миллионам людей благодаря разнообразной издательской программе, которая включает научные работы по всем академическим дисциплинам, библии, музыку, школьные и университетские учебники, книги по бизнесу, словари и справочники, а также академические журналы.
Watch: Как листья сообщают друг другу об атаке жучка
Поделиться
Статья
Вы можете поделиться этой статьей с указанием авторства 4.0 Международная лицензия.
Новое исследование показывает, как системы связи растений реагируют на угрозы голодных существ.
Результаты показывают, что после ранения растения используют сигналы кальция для предупреждения отдаленных тканей о будущих атаках.
На одном видео вы можете увидеть голодную гусеницу, которая сначала обходит края листа, приближается к основанию листа и одним последним укусом отрывает его от остальной части растения.В течение нескольких секунд вспышка флуоресцентного света охватывает другие листья, сигнал о том, что они должны подготовиться к надвигающейся атаке гусеницы или ее родственников.
Гусеница ест лист, поскольку растение посылает сигнал предупреждения другим листьям. (Фото: Toyota / Gilroy)Этот флуоресцентный свет отслеживает кальций, когда он проникает через ткани растения, обеспечивая электрический и химический сигнал об угрозе. Видео является частью исследования, в котором показано, как глутамат — нейромедиатор в большом количестве у животных — активирует эту волну кальция при повреждении растения.
«На протяжении десятилетий было известно, что повреждение листьев, вызванное механическим ранением или жеванием гусеницы, быстро активирует защитные реакции в отдаленных, неповрежденных листьях растения», — говорит Грегг Хоу, профессор Фонда Университета штата Мичиган. «Но что вызывает такую быструю реакцию, остается загадкой».
Опасно, опасность!
Когда лист ранен, электрический заряд проходит через растение, чтобы предупредить другие ткани о возможной опасности. Но что вызвало этот электрический заряд и как он перемещался по всему предприятию, было неизвестно.
Кальций — один кандидат. Он повсеместно встречается в клетках и часто служит сигналом об изменении окружающей среды. А поскольку кальций несет заряд, он может производить электрический сигнал. Но его трудно отследить, потому что уровень его концентрации быстро растет и падает.
Исследователи создали метод, позволяющий видеть кальций в реальном времени. Они разработали растения, которые производят белок, который флуоресцирует вокруг кальция, что позволяет исследователям отслеживать его присутствие и концентрацию. Затем последовали укусы гусениц, порезы ножницами и сокрушительные раны.
В ответ на каждый вид повреждения растения загораются, поскольку кальций течет с места повреждения на другие листья. Сигнал двигался быстро, примерно один миллиметр в секунду, достигнув отдаленных листьев всего за пару минут.
Через несколько минут уровень защитного гормона, жасмоната, в этих далеких листьях резко увеличился. Они готовили завод к будущим угрозам, производя ядовитые химические вещества, отпугивающие хищников.
Соединение множества точек
Предыдущее исследование швейцарского ученого Теда Фармера продемонстрировало, что электрические сигналы, связанные с защитой, зависят от рецепторов глутамата, аминокислоты и основного нейромедиатора, который способствует обмену информацией на большие расстояния между животными и растениями.Фармер показал, что мутантные растения, у которых отсутствуют рецепторы глутамата, теряют свои электрические ответы на угрозы.
После этого исследования исследователи протестировали мутанты, которые нокаутируют рецепторы глутамата. Кальций почти не проявлялся на видео в виде крайних вспышек света.
Результаты показывают, что выход глутамата из раны растения приводит к быстрому распространению кальциевой волны, что, в свою очередь, приводит к выработке жасмоната и защитных реакций.
Исследование объединяет десятилетия исследований, которые показали, как растения выработали умные защитные стратегии в отсутствие центральной нервной системы.Это исследование позволяет лучше всего взглянуть на эти системы связи на предприятиях, которые обычно скрыты от глаз.
«Мы часто думаем, что растения пассивны и находятся во власти окружающей среды. У меня буквально отвисла челюсть, когда я впервые увидел эти видео из лаборатории Гилроя — они прекрасно иллюстрируют, насколько на самом деле активные и сложные растения », — говорит Хоу.
Масацугу Тойота руководил работой в качестве постдокторанта в лаборатории Саймона Гилроя в Университете Висконсин-Мэдисон.Гилрой и Toyota сотрудничали с Хоу и исследователями из Японского агентства науки и технологий и Университета Миссури.
Национальный научный фонд, Министерство энергетики, НАСА и Японское агентство научных технологий PRESTO, KAKENHI, финансировали исследование.
Источник: Университет штата Мичиган
.